Салат листовой: полезные свойства и калорийность
Салат, он же салат зеленый или листовой (Lactuca sativa L.) — травянистое растение семейства Астровые, ценная овощная культура. Питательные и полезные свойства салата листового были известны еще в древнем Риме. Салат был одним из популярнейших продуктов питания на Юге Европы — его употребляли с оливковым маслом и с солью, часто как гарнир к мясу и рыбе.
В наше время культивируется как овощное растение по всем миру. Различают более сотни различных видов — они различаются по форме и цвету листьев, вкусовым качествам, условиям произрастания. Делятся в основном на кочанные и листовые сорта листового салата. И практически все сорта салата зеленого являются полезным и вкусным продуктом питания.
Химический состав
Огромная польза салата листового как овощной культуры объясняется его богатейшим химическим составом. Листья растения состоят из воды, клетчатки, углеводов, незначительного количества белка и жиров.
Состав зеленого салата содержит массу полезных элементов: железо, фосфор, йод, кобальт, цинк, натрий, калий, медь.
Также салат листовой, состав которого содержит очень много каротина и каротиноидов, благотворно влияет на зрение. Полезные свойства салата зеленого также в том, что благодаря высокому содержанию в составе фолиевой кислоты, он совершенно необходим беременным и кормящим женщинам, а также детям для нормального формирования скелета и развития костей.
Польза
Специалисты считают, что регулярное употребление салата в пищу благодаря его полезным свойствам способно предотвратить развитие болезни Альцгеймера, укрепить скелет и костную ткань, наладить пищеварительный процесс за счет большого содержания клетчатки, улучшить состояние зубов, кожи, слизистых оболочек организма, восполнить дефицит витаминов и минеральных элементов.
Калорийность
Салат листовой калорийность которого очень низкая — всего 15 калорий на 100 граммов зеленых листьев, — это идеальный диетический продукт. Ему приписывают способность снижать уровень сахара в крови и противостоять ожирению. За счет высокого содержания аскорбиновой кислоты зеленый листовой салат является прекрасным антиоксидантом и противостоит старению клеток.
Ему приписывают способность снижать уровень сахара в крови и противостоять ожирению. За счет высокого содержания аскорбиновой кислоты зеленый листовой салат является прекрасным антиоксидантом и противостоит старению клеток.
Обычно его употребляют в пищу в свежем виде — в основном в составе витаминных салатов, свежих закусок, гарниров к различным блюдам.
Салат-латук — описание, состав, калорийность и пищевая ценность
15 килокалорий |
Салат латук (латук посевной) — в кулинарии листья молодого одноименного растения, популярной сельскохозяйственной культуры, выращиваемой по всему миру, в регионах с субтропическим и умеренным климатом. Отличаются более нежной, чем у взрослых растений структурой мякоти и менее горьким вкусом.
Существует несколько видов посевного латука, относящихся к двум основным разновидностями данного растения — листовые и кочанные (листья образуют кочан или полукочан различной плотности округлой либо шаровидной формы).
В 100 граммах посевного латука содержится около 15 ккал.
СоставХимический состав салата латук характеризуется повышенным содержанием белков, углеводов, клетчатки, витаминов (A, B4, B9, C, K), макро- (калий, кальций, магний, натрий, фосфор) и микроэлементов (марганец, медь).
Как подавать и готовитьБлагодаря нежной мякоти и приятному кисло-сладкому вкусу, посевной латук нашел очень широкое применение в кулинарии. Его листья не только добавляются в овощные салаты, но и используются при приготовлении огромного количества блюд из мяса, домашней птицы, рыбы и морепродуктов. При этом латук применяется, как вкусовая и витаминизированная добавка, так и декоративное украшение. В этой связи нельзя не упомянуть о необходимости избегать чрезмерной термической обработки этой листовой зелени, при которой она теряет значительную часть своей первоначальной пищевой ценности.
Наиболее качественным считается салат латук с упругими хрустящими листьями без каких-либо повреждений.
Листовой салат латук следует хранить в холодильнике, употребив его в пищу в течение нескольких дней. Куда лучше сохраняются кочанные разновидности этой листовой зелени. При температуре около 10 градусов по Цельсию, хорошей вентиляции и вдали от источников тепла или холода их можно хранить на протяжении 4 недель.
Полезные свойстваСочетание низкой калорийности и богатого жизненно необходимыми минералами и витаминами химического состава делает салат латук отличным вариантом для организации диетического питания. Употребление его листьев снижает нервную возбудимость, уменьшает уровень содержания в крови холестерина, стимулирует процессы кроветворения, оказывает иммуностимулирующее, антиоксидантное, противовоспалительное, желчегонное и мочегонное воздействие.
Индивидуальная непереносимость, подагра, хронические колиты и энтероколиты в период обострения, мочекаменная болезнь.
Салат-латук: состав, калорийность и пищевая ценность на 100 г
|
Общая информация
Вода 94,98 г
Энергетическая ценность 15 ккал
Энергия 62 кДж
Белки 1,36 г
Жиры 0,15 г
Неорганические вещества 0,62 г
Углеводы 2,87 г
Клетчатка 1,3 г
Сахар, всего 0,78 г
Углеводы
Глюкоза (декстроза) 0,36 г
Фруктоза 0,43 г
Минералы
Кальций, Ca 36 мг
Железо, Fe 0,86 мг
Магний, Mg 13 мг
Фосфор, P 29 мг
Калий, K 194 мг
Натрий, Na 28 мг
Цинк, Zn 0,18 мг
Медь, Cu 0,029 мг
Марганец, Mn 0,25 мг
Селен, Se 0,6 мкг
Витамины
Витамин С 9,2 мг
Тиамин 0,07 мг
Рибофлавин 0,08 мг
Никотиновая кислота 0,375 мг
Пантотеновая кислота 0,134 мг
Витамин B-6 0,09 мг
Фолаты, всего 38 мкг
Фолиевая кислота, пищевая 38 мкг
Фолиевая кислота, DFE 38 мкг
Холин, всего 13,6 мг
Бетаин 0,2 мг
Витамин A, RAE 370 мкг
Каротин, бета- 4443 мкг
Витамин A, IU 7405 МЕ
Лютеин + зеаксантин 1730 мкг
Витамин Е (альфа-токоферол) 0,22 мг
Токоферол, гамма 0,41 мг
Токоферол, дельта 0,02 мг
Витамин К (филлохинон) 126,3 мкг
Липиды
Жирные кислоты, насыщенные 0,02 г
16:0 0,018 г
18:0 0,002 г
Жирные кислоты, мононенасыщенные 0,006 г
16:1 недифференцированно 0,002 г
18:1 недифференцированно 0,005 г
Жирные кислоты, полиненасыщенные 0,082 г
18:2 недифференцировано 0,024 г
18:3 недифференцированно 0,058 г
Фитостеролы 38 мг
Аминокислоты
Триптофан 0,009 г
Треонин 0,059 г
Изолейцин 0,084 г
Лейцин 0,079 г
Лизин 0,084 г
Метионин 0,016 г
Цистин 0,016 г
Фенилаланин 0,055 г
Тирозин 0,032 г
Валин 0,07 г
Аргинин 0,071 г
Гистидин 0,022 г
Аланин 0,056 г
Аспарагиновая кислота 0,142 г
Глутаминовая кислота 0,182 г
Глицин 0,057 г
Пролин 0,048 г
Серин 0,039 г
Калорийность Салат микс.
 Химический состав и пищевая ценность.
Химический состав и пищевая ценность.Химический состав и анализ пищевой ценности
Пищевая ценность и химический состав
«Салат микс».В таблице приведено содержание пищевых веществ (калорийности, белков, жиров, углеводов, витаминов и минералов) на 100 грамм съедобной части.
| Нутриент | Количество | Норма** | % от нормы в 100 г | % от нормы в 100 ккал | 100% нормы |
| Калорийность | 16 кКал | 1684 кКал | 6.3% | 10525 г | |
| Белки | 1. 5 г 5 г | 76 г | 2% | 12.5% | 5067 г |
| Жиры | 0.2 г | 56 г | 0.4% | 28000 г | |
| Углеводы | 2 г | 219 г | 0.9% | 5.6% | 10950 г |
Энергетическая ценность Салат микс составляет 16 кКал.
Основной источник: Создан в приложении пользователем. Подробнее.
** В данной таблице указаны средние нормы витаминов и минералов для взрослого человека. Если вы хотите узнать нормы с учетом вашего пола, возраста и других факторов, тогда воспользуйтесь приложением «Мой здоровый рацион».
Салат вечерний – Салат Вечерний рецепт с фото
Рецепт салат «Вечер». Калорийность, химический состав и пищевая ценность.
Пищевая ценность и химический состав «салат «Вечер»».
В таблице приведено содержание пищевых веществ (калорийности, белков, жиров, углеводов, витаминов и минералов) на 100 грамм съедобной части.
| Нутриент | Количество | Норма** | % от нормы в 100 г | % от нормы в 100 ккал | 100% нормы |
| Калорийность | 121.8 кКал | 1684 кКал | 7.2% | 5.9% | 1383 г |
| Белки | 10 г | 76 г | 13.2% | 10. 8% 8% | 760 г |
| Жиры | 8.3 г | 56 г | 14.8% | 12.2% | 675 г |
| Углеводы | 2.4 г | 219 г | 1.1% | 0.9% | 9125 г |
| Органические кислоты | 0.1 г | ~ | |||
| Пищевые волокна | 1 г | 20 г | 5% | 4.1% | 2000 г |
| Вода | 76.7 г | 2273 г | 3.4% | 2.8% | 2963 г |
| Зола | 2.1715 г | ~ | |||
| Витамины | |||||
| Витамин А, РЭ | 65 мкг | 900 мкг | 7.2% | 5.9% | 1385 г |
| бета Каротин | 0.701 мг | 5 мг | 14% | 11.5% | 713 г |
| бета Криптоксантин | 3.5758 мкг | ~ | |||
| Лютеин + Зеаксантин | 1420.2424 мкг | ~ | |||
| Витамин В1, тиамин | 0.066 мг | 1.5 мг | 4.4% | 3.6% | 2273 г |
| Витамин В2, рибофлавин | 0. 132 мг 132 мг | 1.8 мг | 7.3% | 6% | 1364 г |
| Витамин В4, холин | 7.22 мг | 500 мг | 1.4% | 1.1% | 6925 г |
| Витамин В5, пантотеновая | 0.199 мг | 5 мг | 4% | 3.3% | 2513 г |
| Витамин В6, пиридоксин | 0.057 мг | 2 мг | 2.9% | 2.4% | 3509 г |
| Витамин В9, фолаты | 36.636 мкг | 400 мкг | 9.2% | 7.6% | 1092 г |
| Витамин C, аскорбиновая | 9.45 мг | 90 мг | 10.5% | 8.6% | 952 г |
| Витамин Е, альфа токоферол, ТЭ | 1.112 мг | 15 мг | 7.4% | 6.1% | 1349 г |
| бета Токоферол | 0.0003 мг | ~ | |||
| гамма Токоферол | 0.0718 мг | ~ | |||
| дельта Токоферол | 0.0024 мг | ~ | |||
| Витамин Н, биотин | 0.236 мкг | 50 мкг | 0.5% | 0. 4% 4% | 21186 г |
| Витамин К, филлохинон | 60 мкг | 120 мкг | 50% | 41.1% | 200 г |
| Витамин РР, НЭ | 4.122 мг | 20 мг | 20.6% | 16.9% | 485 г |
| Ниацин | 2.239 мг | ~ | |||
| Бетаин | 0.0606 мг | ~ | |||
| Макроэлементы | |||||
| Калий, K | 268.58 мг | 2500 мг | 10.7% | 8.8% | 931 г |
| Кальций, Ca | 73.15 мг | 1000 мг | 7.3% | 6% | 1367 г |
| Магний, Mg | 35.39 мг | 400 мг | 8.8% | 7.2% | 1130 г |
| Натрий, Na | 469.7 мг | 1300 мг | 36.1% | 29.6% | 277 г |
| Сера, S | 5.76 мг | 1000 мг | 0.6% | 0.5% | 17361 г |
| Фосфор, Ph | 111.6 мг | 800 мг | 14% | 11.5% | 717 г |
| Хлор, Cl | 10.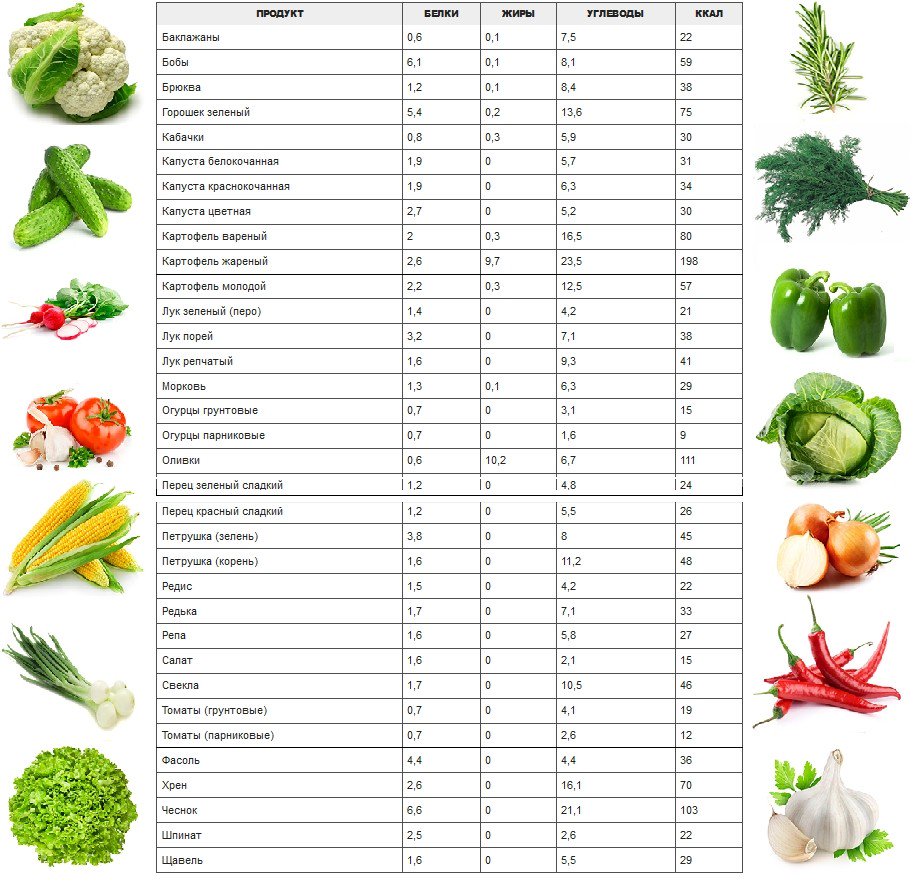 15 мг 15 мг | 2300 мг | 0.4% | 0.3% | 22660 г |
| Микроэлементы | |||||
| Алюминий, Al | 24.2 мкг | ~ | |||
| Бор, B | 29.5 мкг | ~ | |||
| Железо, Fe | 1.97 мг | 18 мг | 10.9% | 8.9% | 914 г |
| Йод, I | 0.48 мкг | 150 мкг | 0.3% | 0.2% | 31250 г |
| Кобальт, Co | 1.212 мкг | 10 мкг | 12.1% | 9.9% | 825 г |
| Марганец, Mn | 0.217 мг | 2 мг | 10.9% | 8.9% | 922 г |
| Медь, Cu | 82.39 мкг | 1000 мкг | 8.2% | 6.7% | 1214 г |
| Молибден, Mo | 1.061 мкг | 70 мкг | 1.5% | 1.2% | 6598 г |
| Никель, Ni | 2.152 мкг | ~ | |||
| Рубидий, Rb | 52 мкг | ~ | |||
| Селен, Se | 1.203 мкг | 55 мкг | 2.2% | 1. 8% 8% | 4572 г |
| Фтор, F | 4.95 мкг | 4000 мкг | 0.1% | 0.1% | 80808 г |
| Хром, Cr | 0.88 мкг | 50 мкг | 1.8% | 1.5% | 5682 г |
| Цинк, Zn | 0.3024 мг | 12 мг | 2.5% | 2.1% | 3968 г |
| Усвояемые углеводы | |||||
| Крахмал и декстрины | 0.071 г | ~ | |||
| Моно- и дисахариды (сахара) | 1.6 г | max 100 г | |||
| Галактоза | 0.0164 г | ~ | |||
| Глюкоза (декстроза) | 0.0939 г | ~ | |||
| Сахароза | 0.4006 г | ~ | |||
| Фруктоза | 0.0794 г | ~ | |||
| Незаменимые аминокислоты | 0.0173 г | ~ | |||
| Аргинин* | 0.0247 г | ~ | |||
| Валин | 0.0151 г | ~ | |||
| Гистидин* | 0.0077 г | ~ | |||
| Изолейцин | 0.0133 г | ~ | |||
| Лейцин | 0. 0238 г 0238 г | ~ | |||
| Лизин | 0.0186 г | ~ | |||
| Метионин | 0.0052 г | ~ | |||
| Метионин + Цистеин | 0.0012 г | ~ | |||
| Треонин | 0.014 г | ~ | |||
| Триптофан | 0.0038 г | ~ | |||
| Фенилаланин | 0.0154 г | ~ | |||
| Фенилаланин+Тирозин | 0.0042 г | ~ | |||
| Заменимые аминокислоты | 0.0402 г | ~ | |||
| Аланин | 0.0167 г | ~ | |||
| Аспарагиновая кислота | 0.0347 г | ~ | |||
| Глицин | 0.017 г | ~ | |||
| Глутаминовая кислота | 0.0528 г | ~ | |||
| Пролин | 0.0193 г | ~ | |||
| Серин | 0.014 г | ~ | |||
| Тирозин | 0.0107 г | ~ | |||
| Цистеин | 0.0048 г | ~ | |||
| Стеролы (стерины) | |||||
| Холестерин | 23.64 мг | max 300 мг | |||
| Фитостеролы | 1.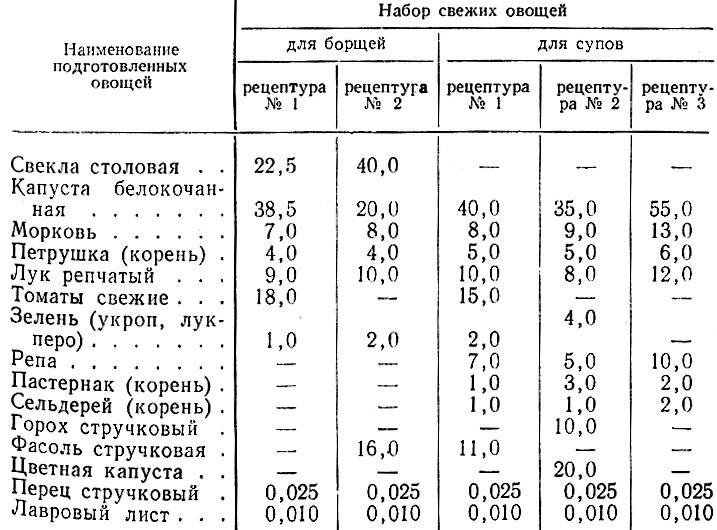 4545 мг 4545 мг | ~ | |||
| бета Ситостерол | 6.0606 мг | ~ | |||
| Жирные кислоты | |||||
| Трансжиры | 0.0004 г | max 1.9 г | |||
| мононенасыщенные трансжиры | 0.0002 г | ~ | |||
| полиненасыщенные трансжиры | 0.0002 г | ~ | |||
| Насыщенные жирные кислоты | |||||
| Насыщеные жирные кислоты | 1.8 г | max 18.7 г | |||
| 12:0 Лауриновая | 0.001 г | ~ | |||
| 14:0 Миристиновая | 0.0002 г | ~ | |||
| 15:0 Пентадекановая | 0.0001 г | ~ | |||
| 16:0 Пальмитиновая | 0.8152 г | ~ | |||
| 17:0 Маргариновая | 0.0001 г | ~ | |||
| 18:0 Стеариновая | 0.1555 г | ~ | |||
| 20:0 Арахиновая | 0.0522 г | ~ | |||
| 22:0 Бегеновая | 0.0006 г | ~ | |||
| 24:0 Лигноцериновая | 0.0003 г | ~ | |||
| Мононенасыщенные жирные кислоты | 4. 1562 г 1562 г | min 16.8 г | 24.7% | 20.3% | |
| 16:1 Пальмитолеиновая | 0.0946 г | ~ | |||
| 16:1 цис | 0.0003 г | ~ | |||
| 18:1 Олеиновая (омега-9) | 3.9825 г | ~ | |||
| 18:1 цис | 0.028 г | ~ | |||
| 18:1 транс | 0.0002 г | ~ | |||
| 20:1 Гадолеиновая (омега-9) | 0.0407 г | ~ | |||
| 22:1 Эруковая (омега-9) | 0.0383 г | ~ | |||
| 22:1 цис | 0.0383 г | ~ | |||
| 24:1 Нервоновая, цис (омега-9) | 0.0025 г | ~ | |||
| Полиненасыщенные жирные кислоты | 0.8917 г | от 11.2 до 20.6 г | 8% | 6.6% | |
| 18:2 Линолевая | 0.7879 г | ~ | |||
| 18:2 транс-изомер, не определён | 0.0002 г | ~ | |||
| 18:2 Омега-6, цис, цис | 0.0132 г | ~ | |||
| 18:3 Линоленовая | 0.0901 г | ~ | |||
| 18:3 Омега-3, альфа-линоленовая | 0. 0139 г 0139 г | ~ | |||
| 20:2 Эйкозадиеновая, Омега-6, цис, цис | 0.0004 г | ~ | |||
| 20:3 Эйкозатриеновая | 0.0009 г | ~ | |||
| 20:3 Омега-3 | 0.0009 г | ~ | |||
| 20:4 Арахидоновая | 0.0007 г | ~ | |||
| Омега-3 жирные кислоты | 0.1 г | от 0.9 до 3.7 г | 11.1% | 9.1% | |
| Омега-6 жирные кислоты | 0.8 г | от 4.7 до 16.8 г | 17% | 14% |
Энергетическая ценность салат «Вечер» составляет 121,8 кКал.
Основной источник: Создан в приложении пользователем. Подробнее.
** В данной таблице указаны средние нормы витаминов и минералов для взрослого человека. Если вы хотите узнать нормы с учетом вашего пола, возраста и других факторов, тогда воспользуйтесь приложением «Мой здоровый рацион».
Салат «Вечерний каприз». Идеальный салат для романтического ужина!
Это ошеломляюще вкусный салат, который удивит вас изысканным сочетанием вкусов и своей необычностью. Он такой вкусный, лёгкий и красивый! А что ещё нужно, чтобы украсить ужин, тем более — ужин романтический? Такой салат не сделает ваш желудок тяжелым, а вас — ленивым.
Он такой вкусный, лёгкий и красивый! А что ещё нужно, чтобы украсить ужин, тем более — ужин романтический? Такой салат не сделает ваш желудок тяжелым, а вас — ленивым.
Такой салат освежает, немного бодрит и наилучшим образом сочетается с бокалом красного вина. Ну, а уж если хочется чего-то более серьезного, то этот салат будет прекрасным гарниром к стейку из говядины.
Ингредиенты на 2 порции Для заправки: + 1 ст. ложка дижонской горчицы (обычной или зернистой), + 1 ст. ложка яблочного уксуса, + 1 ст. ложка мёда, + 4 ст. ложки оливкового масла, + 1/2 ч. ложки мелко порезанного чеснока, + соль и перец по вкусу.
Для салата: + 2 чашки листьев шпината (если листья очень крупные — порежьте их), + 2 чашки листьев салата (можете взять любой микс), + 1/4 чашки мягкого козьего сыра (если нет козьего, то допустимо заменить его на фету), + 1/4 чашки арахиса в сахаре, + 1/4 чашки сушеной канадской клюквы (допускается заменить на кисло-сладкий изюм), + 1 груша.
Как приготовить
Заправка Смешайте все ингредиенты, так чтобы масса стала однородной. Можно воспользоваться блендером.
Примечание: Такая заправка подходит ко многим салатам из свежих листиков, овощей и фруктов. Поэтому, если у вас осталась заправка, просто уберите ее в холодильник и добавляйте в свои любимые салаты. Готовая заправка хранится в холодильнике в стеклянной банке под крышкой до 2 недель.
Салат 1. Промыть и просушить листья шпината и салата. 2. Грушу помыть и нарезать тонкими ломтиками. 3. Выложить на тарелку листья шпината, сверху листья салата. 4. Сверху равномерно распределить ломтики груши, орехи в сахаре и сушеную клюкву. 5. Сверху добавьте заправку, количество определите по вашему усмотрению, но не менее 4-5 ст. ложек. 6. Сверху покрошите кусочки козьего сыра.
Внимание! Такой салат нужно сервировать и заправлять непосредственно перед подачей на стол, чтобы он не утратил свою красоту и свежесть.
Вы можете сервировать его как в общей тарелке, так и сразу в индивидуальных. Заправку можно подать к столу дополнительно, чтобы при желании каждый мог добавить себе желаемое количество соуса.
Это очень вкусно! Попробуйте.
Видео: Женский Интерес
Новые видео!
Не пропустите наши премьеры на YouTube!
Подписывайтесь и смотрите!
Подписаться
- ВЕЧЕРНИЙ САЛАТ ДЛЯ ПОХУДЕНИЯ. МИНУС 5 КИЛО ЗА 2 НЕДЕЛИ! ВЕЧЕРНИЙ САЛАТ ДЛЯ ПОХУДЕНИЯ. МИНУС 5 КИЛО ЗА 2 НЕДЕЛИ! 2 474 просмотра ⌚10 месяцев назад
- ВСЕ МОИ АУДИОКНИГИ = https://www.youtube.com/playlist?list=PLpYgFtDH82dNXMY9v3… Группа вк … ВЕЧЕРНИЙ ЖОР | САЛАТ ПОМИДОРЫ КАБАЧКИ КАХЕТЫ | ВЫЖИТЬ В ГОРОДЕ ВЛОГ 641 просмотр ⌚10 месяцев назад
- Всем привет! Сегодня вас ждет подборка из пяти салатов которые отлично подойдут на ужин. Каждый из них и… 5 Диетических Салатов Вместо Ужина. Ешь вечером — все равно похудеешь! 9 981 просмотр ⌚10 месяцев назад
- Сегодня вы узнаете рецепт белкового салата на вечер, который отлично подойдет для худеющих.
 В этом салате… БЕЛКОВЫЙ САЛАТ НА ВЕЧЕР (Для худеющих) 432 ⌚11 месяцев назад
В этом салате… БЕЛКОВЫЙ САЛАТ НА ВЕЧЕР (Для худеющих) 432 ⌚11 месяцев назад - Салат Вечерний Лучший рецепт при похудении Салат Ем и худею Похудела на 31 кг Салат Вечерний:… Салат Вечерний Лучший рецепт при похудении Салат Ем и худею Похудела на 31 кг 31 721 просмотр ⌚11 месяцев назад
- САЛАТ КОТОРЫЙ ЕДЯТ ГОЛЛИВУДСКИЕ КРАСОТКИ ✿ Вечерний салат ДЛЯ ПОХУДЕНИЯ ✿ ПП САЛАТКак похудеть быстро,… САЛАТ КОТОРЫЙ ЕДЯТ ГОЛЛИВУДСКИЕ КРАСОТКИ
 В этом салате… БЕЛКОВЫЙ САЛАТ НА ВЕЧЕР (Для худеющих) 432 ⌚11 месяцев назад
В этом салате… БЕЛКОВЫЙ САЛАТ НА ВЕЧЕР (Для худеющих) 432 ⌚11 месяцев назадПотери при тепловой и холодной обработке, химический состав: Зелень Салат
Единица измерения: кг Масса единицы измерения: 1000 грамм Средняя цена в РФ за единицу измерения 400 ₽
Виды обработок, проценты потерь массы и пищевых веществ продукта «Зелень Салат»
| Обработка* | Потери при холодной обработке: | Потери при тепловой обработке: | Потери после тепловой обработки: | Потери белков | Потери жиров | Потери углеводов |
|---|---|---|---|---|---|---|
| Листовой холодная обработка | 28,00 | 0,00 | 0,00 | 0,00 | 0,00 | 0,00 |
| Кочанный холодная обработка | 33,00 | 0,00 | 0,00 | 0,00 | 0,00 | 0,00 |
Зелень Салат, и еще 1100+ продуктов есть в «Шеф Эксперт»! Подробнее. ..
..
Зелень Салат — Химический состав
| Белки (гр/100 гр продукта): | 1,50 |
| Жиры (гр/100 гр продукта): | 0,20 |
| Углеводы (гр/100 гр продукта): | 2,30 |
| Сухие вещества (гр/100 гр продукта): | 6,00 |
| Влажность в %: | 94,00 |
| Калорийность, ккал: | 17,00 |
| Калорийность, кДж: | 71,18 |
Информация для составления технологической карты (массовые доли)
Эти данные потребуются при оформлении Технико-технологической карты на блюда, в состав которых входит Зелень Салат. Для расчета массовых долей жира и сахара необходимо знать, содержит ли продукт так называемый «свободный» жир и сахар (т.е., определяемый при лабораторном анализе).
| Содержит свободный жир | Нет |
| Содержит свободный сахар | Нет |
| Содержит спирт | Нет |
Хотите разрабатывать правильные документы для общепита? — скачайте программу «Шеф Эксперт»!
Аллергены в продукте «Зелень Салат»
В соответствии с требованиями Технического регламента ТР ТС 022/2011, при разработке технологической документации на блюда (изделия) указываются аллергены, входящие в состав продукта «Зелень Салат»:
Отсутствуют аллергены из перечня технического регламента.
Информация об аллергенах в блюде необходима также для контроля в соответствие с принципами ХАССП.
Комментарии:
- Данные о пищевой ценности продукта приведены из источников, рекомендованных к применению Федеральной службой по Надзору в сфере защиты прав потребителей и благополучия человека (Роспотребнадзор)
Как рассчитать потери и калорийность блюд:
Расчет расхода сырья, калорийность, выход и себестоимость блюд должен уметь делать любой шеф-повар или су-шеф.
Проблема в том, что для расчета технико-технологической карты и калькуляций по всем требованиям, нужны опыт и время.
Программа Шеф Эксперт позволяет разработать весь комплект документов на блюдо за несколько минут. При этом:
— Не нужны справочники химического состава и сборники рецептур, т.к. все данные уже есть в программе
— Не требуются знания технолога. В программе достаточно подобрать ингредиенты, входящие в состав блюда и указать их массу. Все остальные расчеты будут выполнены автоматически.
Все остальные расчеты будут выполнены автоматически.
Скачайте демо-версию, и пользуйтесь бесплатно 30 дней…
* Мы тщательно следим за достоверностью данных в справочнике химсостава. Но реальные потери и состав продуктов могут отличаться от указанных здесь, в-зависимости от сезонных и других факторов.
Калорийность крабовая закуска. Калорийность Крабовый салат. Химический состав и пищевая ценность. Салат «Крабовый с рисом»
Крабовый салат занимает одно из лидирующих мест на нашем праздничном столе в наше время. На сегодняшний день можно с большой уверенностью сказать, что это блюдо одно их самых популярных из всех имеющихся. Сколько у крабового салата из крабовых палочек калорийность — вопрос этот не может оставить многих людей равнодушными. Многих интересует вопрос – насколько могут быть полезен такой продукт, как крабовые палочки? На все волнующие вопросы постараюсь ответить в дальнейшем рассмотрении данного рецепта.
Какова польза крабового салата?
Давайте с вами уточним тот факт, что крабовые палочки, которые мы видим в магазинах – это не искусственный продукт, так как они в основном состоят из концентрированного рыбного белка. Рекомендую брать натуральное крабовое мясо, оно полезно и хорошо усваивается.
Рекомендую брать натуральное крабовое мясо, оно полезно и хорошо усваивается.
Блюдо очень полезно, особенно для тех, кому не хватает в рационе животного жира, белков, минералов и других различных витаминов, а также тем людям, у которых понижено питание. Достаточно употреблять в день около 100-150 грамм и все расстройства, которые только начинают развиваться на первоначальных стадиях очень быстро исчезнут.
Давайте с Вами разберемся — сколько калорийности в крабовом салате из крабовых палочек содержится.
Калорийность его составляет порядком 73 калорий на 100 грамм продукта. Как правило, в классическом рецепте, калорийность блюда составляет примерно 153 калорий на 100 грамм продукта. Калории крабового салата могут варьироваться, в зависимости от содержания рецепта.
Давайте с Вами и приготовим один из его вариантов. Для этого нам понадобится:
Ингредиенты на 4 порции:
1 банка варёной кукурузы;
3 куриных яйца;
1/3 стакана риса;
250 грамм крабовых палочек;
¼ средней головки репчатого лука;
майонез;
соль;
зелень по вкусу.
Для начала, необходимо хорошо промыть рис, затем бросить его в 1 литр кипящей воды. Варить следует примерно 20 минут, периодически помешивая. Следите за тем, чтобы рис не разварился, иначе блюдо в целом получится не таким, каким хочу Вам преподнести. Советую, добавить в конце варки риса одну столовую ложку лимонного сока и рис ваш станет белоснежным. Промойте рис в теплой кипяченой воде и дайте ему остыть.
Следующим действием, отварить яйца вкрутую, после чего остужайте и очищайте от скорлупы. Резать яйца необходимо в виде мелких кубиков вручную или при помощи яйцерезки. Крабовые палочки резать следует также мелкими кубиками. Нарезайте половину репчатого лука и хорошенько измельчайте. Лук можно класть в блюдо по Вашему желанию. Открывайте кукурузу и сливайте жидкость, после чего засыпайте в основное глубокое блюдо.
Все обработанные компоненты укладываем в глубокое блюдо, заправляем майонезом и тщательно перемешиваем. Наш крабовый рецепт полностью готов! Кушайте на здоровье! Не бойтесь того, сколько калорийности у крабового салата из крабовых палочек. Желаю Вам приятного аппетита!
Желаю Вам приятного аппетита!
Приготовить его довольно просто, при этом не нужно обладать особыми кулинарными навыками. Большинство продуктов не требует дополнительной тепловой обработки, их достаточно только правильно нарезать. Рецепты салатов из крабовых палочек представлены в различных вариантах и каждый может найти для себя то, что придется по вкусу. В него добавляют зеленый горошек, свежие помидоры и огурцы, зелень, курицу, фасоль, рис, чеснок, и даже ананас. Украсить такое угощение можно тертым сыром, кусочками черри или мелко нарезанной зеленью.
Пять самых часто используемых ингредиентов в рецептах:
Такая красота станет настоящим украшением простого семейного ужина или праздничного стола. Чтобы правильно приготовить блюдо с крабовыми палочками, необходимо внимательно следовать рекомендациям в рецепте и соблюдать указанные пропорции. Это позволит сочетать нужные вкусовые оттенки в равной степени и создать настоящий кулинарный шедевр, который по достоинству оценят и гости, и домашние.
Крабовый салат уже давно стал завсегдатаем на праздничном и повседневном столе во многих домах. Он простой, приятный на вкус, его отлично едят и дети, и взрослые. Если вы следите за своей фигурой, обратите на то, что есть разные рецепты этого блюда, и можно выбрать тот вариант салата из крабовых палочек, калорийность которого будет наиболее низкой.
Рецепты и калорийность крабового салата
Поскольку этот салат многовариантен, стоит по отдельности рассматривать калорийность разных его рецептов. Мs выбрали несколько самых популярных и проанализировали их с точки зрения пищевой ценности. Зная, сколько калорий в крабовом салате, вы сможете легко вписать его в рацион без вреда для фигуры.
Салат «Крабовый с рисом»
Ингредиенты:
- кукуруза сладкая консервированная — 235 г;
- палочки крабовые — 200 г;
- рис отварной — 200 г;
- яблоко — 100 г;
- лук репчатый — 20 г;
- майонез — 100 г;
- яйцо — 240 г.
Приготовление
Нашинкуйте мелко отварные яйца, крабовые палочки, лук и яблоко. Добавьте к этим компонентам кукурузу и предварительно отваренный рис (лучше брать пропаренный или длиннозерный). Заправьте майонезом и дайте настояться 20 минут.
Калорийность крабового салата с майонезом 197.7 ккал, из них 6.2 г приходится на белки, 9.1 г – на жиры, 22.6 г на . Учитывая калорийность и заправку, есть такой салат лучше не позднее обеда.
Салат «Крабовый с рисом»
Ингредиенты:
Приготовление
Нашинкуйте кубиками отварные яйца, крабовые палочки и огурец. Добавьте к этим компонентам кукурузу, заправьте майонезом и дайте настояться 20 минут.
Энергетическая ценность такого салата — 128 ккал, из них 9.2 г — белки, 7.4 г – жиры, 5.9 г — углеводы. Если заменить майонез на белый йогурт, этот вариант вполне сгодится и для ужина.
Крабовый салат богат такими витаминами и минералами, как: витамином А — 22,2 %, холином — 13,7 %, витамином E — 73,3 %, витамином H — 19,8 %, кремнием — 67,7 %, фосфором — 18,7 %, железом — 12,8 %, кобальтом — 39 %, марганцем — 19,7 %, медью — 12 %, молибденом — 15,1 %, селеном — 18,4 %
- Витамин А отвечает за нормальное развитие, репродуктивную функцию, здоровье кожи и глаз, поддержание иммунитета.
- Холин входит в состав лецитина, играет роль в синтезе и обмене фосфолипидов в печени, является источником свободных метильных групп, действует как липотропный фактор.
- Витамин Е обладает антиоксидантными свойствами, необходим для функционирования половых желез, сердечной мышцы, является универсальным стабилизатором клеточных мембран. При дефиците витамина Е наблюдаются гемолиз эритроцитов, неврологические нарушения.
- Витамин Н участвует в синтезе жиров, гликогена, метаболизме аминокислот. Недостаточное потребление этого витамина может вести к нарушению нормального состояния кожных покровов.
- Кремний входит в качестве структурного компонента в состав гликозоаминогликанов и стимулирует синтез коллагена.
- Фосфор принимает участие во многих физиологических процессах, включая энергетический обмен, регулирует кислотно-щелочного баланса, входит в состав фосфолипидов, нуклеотидов и нуклеиновых кислот, необходим для минерализации костей и зубов. Дефицит приводит к анорексии, анемии, рахиту.
- Железо входит в состав различных по своей функции белков, в том числе ферментов. Участвует в транспорте электронов, кислорода, обеспечивает протекание окислительно- восстановительных реакций и активацию перекисного окисления. Недостаточное потребление ведет к гипохромной анемии, миоглобиндефицитной атонии скелетных мышц, повышенной утомляемости, миокардиопатии, атрофическому гастриту.
- Кобальт входит в состав витамина В12. Активирует ферменты обмена жирных кислот и метаболизма фолиевой кислоты.
- Марганец участвует в образовании костной и соединительной ткани, входит в состав ферментов, включающихся в метаболизм аминокислот, углеводов, катехоламинов; необходим для синтеза холестерина и нуклеотидов. Недостаточное потребление сопровождается замедлением роста, нарушениями в репродуктивной системе, повышенной хрупкостью костной ткани, нарушениями углеводного и липидного обмена.
- Медь входит в состав ферментов, обладающих окислительно-восстановительной активностью и участвующих в метаболизме железа, стимулирует усвоение белков и углеводов. Участвует в процессах обеспечения тканей организма человека кислородом. Дефицит проявляется нарушениями формирования сердечно-сосудистой системы и скелета, развитием дисплазии соединительной ткани.
- Молибден является кофактором многих ферментов, обеспечивающих метаболизм серусодержащих аминокислот, пуринов и пиримидинов.
- Селен — эссенциальный элемент антиоксидантной системы защиты организма человека, обладает иммуномодулирующим действием, участвует в регуляции действия тиреоидных гормонов. Дефицит приводит к болезни Кашина-Бека (остеоартроз с множественной деформацией суставов, позвоночника и конечностей), болезни Кешана (эндемическая миокардиопатия), наследственной тромбастении.
 Дефицит приводит к анорексии, анемии, рахиту.
Дефицит приводит к анорексии, анемии, рахиту.Полный справочник самых полезных продуктов вы можете посмотреть в приложении
Калорийность крабового салата с майонезом. Сколько калорий в крабовом салате?
Многие очень любят крабовый салат. Он действительно вкусный, так как в нем содержатся прекрасные ингредиенты. Для многих застолий выбирается именно крабовый салат.
Для многих застолий выбирается именно крабовый салат.
Калорийность составляет в среднем 128 ккал. В зависимости от компонентов этот показатель может меняться.
Есть ли польза?
Крабовые палочки содержат витамин В, который необходим для нормальной работы нервной системы.
Витамин А благоприятно воздействует на зрение. Также там есть витамины C и D. Йод поддерживает иммунную систему, которая будет сопротивляться различным болезням.
Магний, находящийся в них, способствует укреплению сосудов и помогает работе сердца.
Кукуруза обогащена витаминами группы В, С, РР. Она содержит белки и различные микроэлементы. Такой состав благоприятно воздействует на пищеварительную систему и работу головного мозга.
Яйца являются источниками витаминов А, В, D. В свежих огурцах тоже много всего полезного, они очищают организм от токсинов, помогают вывести лишний холестерин. Калорийность крабового салата с майонезом составляет 128 ккал. Как и с любым другим блюдом, с ним необходимо соблюдать меру, так как избыточное количество любой пищи вредно всем. Также это связано с тем, что в блюде находится майонез.
Также это связано с тем, что в блюде находится майонез.
- Калорийность оливье с колбасой и майонезом.
- Калорийность селедки под шубой с майонезом.
Пищевая ценность
Крабовый салат калорийность может иметь разную в зависимости от того, что в него входит. В приготовленном по классическому рецепту блюде: 9,2 г белков; 7,4 г жиров; 5,9 г углеводов.
Часто готовится крабовый салат с рисом. Калорийность такого блюда составляет 197 ккал на 100 г. Чтобы блюдо получилось диетическим, майонез заменяют нежирной сметаной или йогуртом. Вид майонеза тоже влияет на калорийность салата, поэтому важно внимательно выбирать продукт с небольшим процентом жирности. Вместо магазинного можно приготовить домашний соус, который сделает блюдо намного полезным. С такой заправкой получится тоже очень вкусно. Чтобы приготовить салат с рисом, необходимо отварить крупу. Она не должна получиться разваренной. Если в конце варки добавить сок лимона (1 ст. л.), то продукт получится белым. Отварить надо и яйца. Крабовые палочки нарезаются кубиками. Так же обрабатываются яйца. Лук тоже надо нарезать, затем смешать компоненты с кукурузой. Блюдо надо выложить в салатник. По вкусу нужно посолить, заправить майонезом. Перед подачей на стол салат должен постоять в холодильнике. Такое блюдо вкусное и питательное. Рекомендовано его умеренное употребление.
Отварить надо и яйца. Крабовые палочки нарезаются кубиками. Так же обрабатываются яйца. Лук тоже надо нарезать, затем смешать компоненты с кукурузой. Блюдо надо выложить в салатник. По вкусу нужно посолить, заправить майонезом. Перед подачей на стол салат должен постоять в холодильнике. Такое блюдо вкусное и питательное. Рекомендовано его умеренное употребление.
Калорийность крабового салата-150-153 калорий на 100 грамм салата.
Крабовый салат крабовому салату рознь. И калорийность каждого из вариантов разная. Давайте рассмотрим один из классических вариантов.
- Состав: крабовые палочки, куриное яйцо, кукуруза консервированная, круглый белый рис, лук репчатый, майонез, соль и перец по вкусу.
- Такой вариант будет весить примерно 153 калории на 100 граммов готового салата.
- Но можно уменьшить его калорийность, если, к примеру, заменить рис на китайскую капусту.
- Также калорийность уменьшится, если заправлять салат не чистым майонезом, а смешать его 50 на 50 с 10-процентной сметаной.
Крабовый салат — частый гость на праздничном столе и не только. Из-за простоты в приготовлении, вкуса, аппетитного вида крабовый салат завоевал признание многих. В классическом салате обязательно содержатся яйца, крабовые палочки, кукуруза, а кроме того рис, капуста, свежие огурцы, майонез, соль, зелень по вкусу. От того какие ингредиенты выбраны для салата и зависит его калорийность.
В 100 г. салата, содержащего помимо основных ингредиентов еще и свежие огурцы содержится около 130 Ккал .
В салате, где содержится рис — около 197 Ккал .
Еще важный момент: если заправлять блюдо не майонезом, а простым несладким йогуртом, то его калорийность будет ниже.
Виды салата
Поскольку этот салат многовариантен, стоит по отдельности рассматривать калорийность разных его рецептов. Мs выбрали несколько самых популярных и проанализировали их с точки зрения пищевой ценности. Зная, сколько калорий в крабовом салате, вы сможете легко вписать его в рацион без вреда для фигуры.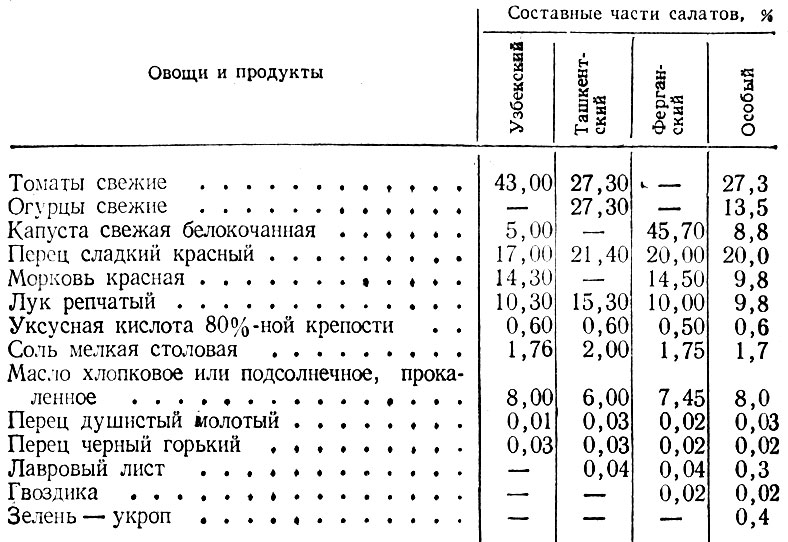
Рецепт и калорийность крабового салата с рисом
Ингредиенты:
- палочки крабовые — 200 г;
- рис отварной — 200 г;
- яблоко — 100 г;
- лук репчатый — 20 г;
- майонез — 100 г;
- яйцо — 240 г.
Приготовление
Нашинкуйте мелко отварные яйца, крабовые палочки, лук и яблоко. Добавьте к этим компонентам кукурузу и предварительно отваренный рис (лучше брать пропаренный или длиннозерный). Заправьте майонезом и дайте настояться 20 минут.
Калорийность крабового салата с майонезом 197.7 ккал, из них 6.2 г приходится на белки, 9.1 г – на жиры, 22.6 г на углеводы. Учитывая калорийность и заправку, есть такой салат лучше не позднее обеда.
Рецепт и калорийность крабового салата без риса с майонезом
Ингредиенты:
- кукуруза сладкая консервированная — 235 г;
- палочки крабовые — 200 г;
- яйцо – 3 шт.;
- огурец – 1 шт.;
- майонез — 100 г.
Приготовление
Нашинкуйте кубиками отварные яйца, крабовые палочки и огурец. Добавьте к этим компонентам кукурузу, заправьте майонезом и дайте настояться 20 минут.
Добавьте к этим компонентам кукурузу, заправьте майонезом и дайте настояться 20 минут.
Энергетическая ценность такого салата — 128 ккал, из них 9.2 г — белки, 7.4 г – жиры, 5.9 г — углеводы. Если заменить майонез на белый йогурт, этот вариант вполне сгодится и для ужина.
Вред салата
Хоть крабовый салат калорийность имеет вполне приемлемую, все же он вреден для организма. В продукте есть химические компоненты (консерванты, красители), например, Е171, Е420 и Е160. Больший вред будет для детей. Поэтому часто употреблять такое блюдо не стоит. В мясе сурими есть искусственные вещества, от которых нет пользы для организма. Если часто питаться таким салатом, то это может стать причиной нарушения работы желудка. Специалисты считают, что употребление всего двух крабовых палочек каждый день может нанести вред пищеварению. Поэтому важно в меру кушать крабовый салат. Калорийность его может навредить фигуре, если не контролировать размер порции.
Салат латук — калорийность, химический состав, гликемический индекс
Содержание пищевых веществ в таблице приведено на 100 грамм продукта.
Калорийность и макронутриенты
Гликемический индекс
Гликемический индекс
Омега 3,6,9
Альфа-линоленовая к-та (18:3) (Омега-3), г
Клетчатка, Холестерин, Трансжиры
Холестерин, мг
Витамины
Альфа-каротин, мкг
Бета-каротин, мкг
Витамин D, кальциферол, мкг
Витамин E, альфа токоферол, мг
Витамин K, филлохинон, мкг
Витамин C, аскорбиновая, мг
Витамин B1, тиамин, мг
Витамин B2, рибофлавин, мг
Витамин B3, витамин PP, ниацин, мг
Витамин B4, холин, мг
Витамин B5, пантотеновая, мг
Витамин B6, пиридоксин, мг
Витамин B7, биотин, мг
Витамин B8, инозит, мг
Витамин B9, фолаты, мкг
Витамин B11, L-карнитин, мг
Витамин B12, кобаламин, мкг
Витамин B13, оротовая кислота, мг
Коэнзим Q10, убихинон, мг
Витамин N, липоевая кислота, мг
Витамин U, метилмегионин-сульфоний, мг
Микроэлементы
Аминокислотный состав
— незаменимые аминокислоты
Фенилаланин, г
Аспарагиновая, г
Глутаминовая, г
Свойства салата — Ботанический онлайн
В этом разделе вы найдете информацию о файлах cookie, которые могут быть созданы с помощью этого веб-сервиса. Botanical-online, как и большинство других веб-сайтов в Интернете, использует свои собственные и сторонние файлы cookie, чтобы улучшить взаимодействие с пользователем и предложить доступный и адаптированный просмотр. Ниже вы найдете подробную информацию о файлах cookie, типах файлов cookie, используемых на этом веб-сайте, о том, как отключить их в вашем браузере и как заблокировать их во время просмотра, таким образом, соблюдение нормативных требований в отношении файлов cookie (Закон 34/2002 г. 11 июля об услугах информационного общества и электронной коммерции (LSSI), который переносит Директиву 2009/136 / CE, также называемую «Директивой о файлах cookie», в испанское законодательство).
Botanical-online, как и большинство других веб-сайтов в Интернете, использует свои собственные и сторонние файлы cookie, чтобы улучшить взаимодействие с пользователем и предложить доступный и адаптированный просмотр. Ниже вы найдете подробную информацию о файлах cookie, типах файлов cookie, используемых на этом веб-сайте, о том, как отключить их в вашем браузере и как заблокировать их во время просмотра, таким образом, соблюдение нормативных требований в отношении файлов cookie (Закон 34/2002 г. 11 июля об услугах информационного общества и электронной коммерции (LSSI), который переносит Директиву 2009/136 / CE, также называемую «Директивой о файлах cookie», в испанское законодательство).
Что такое файлы cookie?
Файлы cookie — это текстовые файлы, которые браузеры или устройства создают при посещении веб-сайтов в Интернете. Они используются для хранения информации о посещении и соответствуют следующим требованиям:
- Для обеспечения правильной работы веб-сайта.
- Установить уровни защиты пользователей от кибератак.
- Для сохранения предпочтений просмотра.
- Чтобы узнать опыт просмотра пользователем
- Для сбора анонимной статистической информации для повышения качества.
- Предлагать персонализированный рекламный контент

Файлы cookie связаны только с анонимным пользователем. Компьютер или устройство не содержат ссылок, раскрывающих личные данные. В любое время можно получить доступ к настройкам браузера, чтобы изменить и / или заблокировать установку отправленных файлов cookie, не препятствуя доступу к контенту. Однако сообщается, что это может повлиять на качество работы служб.
Какую информацию хранит файл cookie?
Файлы cookie обычно не хранят конфиденциальную информацию о человеке, такую как кредитные карты, банковские реквизиты, фотографии, личную информацию и т. Д.Данные, которые они хранят, носят технический характер.
Какие типы файлов cookie существуют?
Существует 2 типа файлов cookie в зависимости от их управления:
- Собственные файлы cookie: те, которые отправляются в браузер или устройство и управляются исключительно нами для наилучшего функционирования веб-сайта.
- Сторонние файлы cookie: те, которые отправляются в браузер или на устройство и управляются третьими сторонами. Они созданы не в нашем домене. У нас нет доступа к сохраненным данным (например, нажатием кнопок социальных сетей или просмотром видео, размещенных на другом веб-сайте), которые устанавливаются другим доменом нашего веб-сайта.Мы не можем получить доступ к данным, хранящимся в файлах cookie других веб-сайтов, когда вы просматриваете вышеупомянутые веб-сайты.

Какие файлы cookie используются на этом веб-сайте?
При просмотре Botanical-online будут созданы собственные и сторонние файлы cookie. Они используются для хранения и управления информацией о конфигурации навигации, веб-аналитики и персонализации рекламы. Сохраненные данные являются техническими и ни в коем случае не личной информацией для идентификации навигатора.
Ниже приведена таблица с указанием наиболее важных файлов cookie, используемых на этом веб-сайте, и их назначения:
Собственные файлы cookie
| Имя файла cookie | Назначение |
| aviso_idioma | Принятие раздела уведомление (язык согласно браузеру посетителя). Технические файлы cookie. Технические файлы cookie. |
| tocplus_hidetoc | Отображение или сбор содержания. Технические файлы cookie |
| adGzcDpEokBbCn XztAIvbJNxM sdLtvFO | Создает случайные буквенно-цифровые данные для защиты веб-сайта путем обнаружения и предотвращения вредоносных действий. Технические файлы cookie. |
Сторонние файлы cookie
| Имя файла cookie | Назначение |
| _gid _ga _gat_gtag_ * | Относится к аналитической или статистической функции посещаемости сайта.Идентификаторы хранятся для подсчета количества посещений, дат доступа, географического положения, а также других статистических функций. Аналитический cookie. |
| __gads | Относится к рекламе, отображаемой на сайте. Рекламный файл cookie Рекламный файл cookie |
| IDE DSID СОГЛАСИЕ NID | Создано службами Google (например, reCaptcha, Youtube, поиск. Технические файлы cookie. |
| Youtube | Файлы cookie для интеграции видеосервиса YouTube на веб-сайт.Социальный файл cookie. |
Как изменить настройки файлов cookie?
Вы можете ограничить, заблокировать или удалить файлы cookie Botanical-online или любой другой веб-сайт, используя свой интернет-браузер. У каждого браузера своя конфигурация. Вы можете увидеть, как действовать дальше, в разделе «Справка». Затем мы показываем список для работы с основными текущими браузерами:
Как изменить настройки файлов cookie на этом сайте?
Напоминаем, что вы можете в любое время просмотреть предпочтения относительно принятия или отказа от файлов cookie на этом сайте, щелкнув «Дополнительная информация» в сообщении о принятии или щелкнув «Политика использования файлов cookie», постоянно присутствующая на всех страницах. сайта.
сайта.
Этот сайт использует файлы cookie для повышения производительности. Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Есть много причин, по которым cookie не может быть установлен правильно. Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie. Вам необходимо сбросить настройки своего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались. Чтобы принять файлы cookie с этого сайта, используйте кнопку «Назад» и примите файлы cookie.
- Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом. Если часы вашего компьютера показывают дату до 1 января 1970 г. ,
браузер автоматически забудет файл cookie. Чтобы исправить это, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie. Вы должны отключить приложение при входе в систему или уточнить у системного администратора.
 ,
браузер автоматически забудет файл cookie. Чтобы исправить это, установите правильное время и дату на своем компьютере.
,
браузер автоматически забудет файл cookie. Чтобы исправить это, установите правильное время и дату на своем компьютере.Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу. Чтобы предоставить доступ без файлов cookie потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в файле cookie может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта. Например, сайт
не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к
остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
Например, сайт
не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к
остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
химический состав салата
; Росалес М.А.По данным Института медицины, допустимый верхний уровень потребления йода составляет. и И.К. Мы также обнаружили, что обогащение йодом положительно влияет на концентрацию некоторых микро- и микронутриентов. Саженцы помещали в лунки (на расстоянии 25 см друг от друга) плит из пенополистирола, растения поливали в течение одной минуты каждые 5 минут в течение дня с 5:00 до 19:00 и во время (электропроводность) питательного раствора всех обработок всего экспериментов было 1.75 мСм. и pH всех питательных растворов был доведен до 5,70 с использованием 38% азотной кислоты. Программа борьбы с ЙДЗ в Индии — это история успеха общественного здравоохранения: 92% населения потребляют йодированную соль. Минеральный состав (средство на 2018-2019 годы). В этом исследовании изучали, отражается ли генетическое разнообразие среди сортов цветной капусты (белая ‘Xenia’ F1, зеленый ‘Vitaverde’ F1, фиолетовый ‘Graffiti’ F1, оранжевый ‘Sunset’ F1, романеско ‘Celio’ F1) и охлаждение трансплантата в содержании 17 элементы зрелого творога.Химический состав салата по содержанию сухого вещества и питательные качества салата. Раствор MetOH в пробирку и инкубирование еще 15 мин при 60, 4; средства, за которыми следует та же буква, не имеют большого значения. Методы: это перекрестное исследование было проведено среди 223 учащихся женского пола, отобранных путем многоступенчатой кластерной выборки из 12 школ. Получите доступ к научным знаниям из любого места. показывает содержание соединений йода в салате, биоусиленном йодом. показали, что листья салата, обогащенного 5-ISA, характеризуются самым высоким уровнем йода.Дефицит йода по-прежнему является глобальной проблемой? и Т. Химический состав салата-латука (Lactuca sativa L.
Минеральный состав (средство на 2018-2019 годы). В этом исследовании изучали, отражается ли генетическое разнообразие среди сортов цветной капусты (белая ‘Xenia’ F1, зеленый ‘Vitaverde’ F1, фиолетовый ‘Graffiti’ F1, оранжевый ‘Sunset’ F1, романеско ‘Celio’ F1) и охлаждение трансплантата в содержании 17 элементы зрелого творога.Химический состав салата по содержанию сухого вещества и питательные качества салата. Раствор MetOH в пробирку и инкубирование еще 15 мин при 60, 4; средства, за которыми следует та же буква, не имеют большого значения. Методы: это перекрестное исследование было проведено среди 223 учащихся женского пола, отобранных путем многоступенчатой кластерной выборки из 12 школ. Получите доступ к научным знаниям из любого места. показывает содержание соединений йода в салате, биоусиленном йодом. показали, что листья салата, обогащенного 5-ISA, характеризуются самым высоким уровнем йода.Дефицит йода по-прежнему является глобальной проблемой? и Т. Химический состав салата-латука (Lactuca sativa L. ), обогащенного йодом с помощью KIO3, 5-йод- и 3,5-дийодосалициловой кислоты при гидропонном культивировании. к применению I + Se и I + Se + трех концентраций SA. Активность vHPO в культурных растениях, а также поглощение и метаболизм йодосалицилатов в салате еще не изучались.В диетах для похудения рекомендуется употреблять сорта с твердыми листьями, поскольку они медленнее перерабатываются организмом и содержат больше клетчатки. Содержание легкоусвояемых углеводов. Хроматографический анализ проводили при следующих условиях: C; объем вводимой пробы — 1 мл; поток газа-носителя C, выдерживаемый в течение 5 мин, использовали для разделения различных метиловых эфиров жирных кислот; затем температуру повышали каждые 5 до 180, еще 16 мин; затем он поднимался каждые 5 до 220 (удар) на полном спектре сканирования от 40 до 500 м.используется для проведения масс-спектрометрического анализа. Ванадий в основном был обнаружен в корнях, где содержание этого элемента увеличивалось пропорционально его дозе.
), обогащенного йодом с помощью KIO3, 5-йод- и 3,5-дийодосалициловой кислоты при гидропонном культивировании. к применению I + Se и I + Se + трех концентраций SA. Активность vHPO в культурных растениях, а также поглощение и метаболизм йодосалицилатов в салате еще не изучались.В диетах для похудения рекомендуется употреблять сорта с твердыми листьями, поскольку они медленнее перерабатываются организмом и содержат больше клетчатки. Содержание легкоусвояемых углеводов. Хроматографический анализ проводили при следующих условиях: C; объем вводимой пробы — 1 мл; поток газа-носителя C, выдерживаемый в течение 5 мин, использовали для разделения различных метиловых эфиров жирных кислот; затем температуру повышали каждые 5 до 180, еще 16 мин; затем он поднимался каждые 5 до 220 (удар) на полном спектре сканирования от 40 до 500 м.используется для проведения масс-спектрометрического анализа. Ванадий в основном был обнаружен в корнях, где содержание этого элемента увеличивалось пропорционально его дозе. сухое вещество по сравнению с контрольными растениями. Подтверждающие: 1, упоминания: 22 — Химический состав и антиоксидантная способность салата: сравнительное исследование типов обычного размера (Ромейн) и детского размера (Маленький драгоценный камень и Мини Ромейн) — Алисия Лопес, Гарсия-Алонсо Хавьер, Хосе Фенолл, Пилар Хеллин, Пилар Флорес трийодтиронин в форме натриевой соли (T3-Na), трийодтиронина (T3) и йодтирозина.Представленный метод расчета HQ представляет собой только потребление I со свежими листьями салата. важен с точки зрения потенциальных потребителей. Растения выращивали в гидропонной системе NFT (Nutrient Film Technique). Авторы заявляют об отсутствии конфликта интересов. (По польски). приобретение, С.С., А.К. Известно, что дефицит йода является самой серьезной причиной предотвратимых повреждений мозга. мл-1 дейтерированной салициловой кислоты (SA-d4, Sigma-Aldrich). действия в нейронах. Мы делаем вывод, что органические соединения йода, т.е.е., 3,5-diISA и 5-ISA, могут поглощаться корнями растений томата на ранней стадии развития.
сухое вещество по сравнению с контрольными растениями. Подтверждающие: 1, упоминания: 22 — Химический состав и антиоксидантная способность салата: сравнительное исследование типов обычного размера (Ромейн) и детского размера (Маленький драгоценный камень и Мини Ромейн) — Алисия Лопес, Гарсия-Алонсо Хавьер, Хосе Фенолл, Пилар Хеллин, Пилар Флорес трийодтиронин в форме натриевой соли (T3-Na), трийодтиронина (T3) и йодтирозина.Представленный метод расчета HQ представляет собой только потребление I со свежими листьями салата. важен с точки зрения потенциальных потребителей. Растения выращивали в гидропонной системе NFT (Nutrient Film Technique). Авторы заявляют об отсутствии конфликта интересов. (По польски). приобретение, С.С., А.К. Известно, что дефицит йода является самой серьезной причиной предотвратимых повреждений мозга. мл-1 дейтерированной салициловой кислоты (SA-d4, Sigma-Aldrich). действия в нейронах. Мы делаем вывод, что органические соединения йода, т.е.е., 3,5-diISA и 5-ISA, могут поглощаться корнями растений томата на ранней стадии развития. Стебли и листья дикого салата нарезали на более мелкие кусочки, высушили и измельчили в очень мелкий порошок. Миссури, США). (TCDD) вызывает нарушение экспрессии рецептора NMDA в культурах неокортикальных клеток крыс. REDL) «Lollo rossa» и «Redin» с маленькими красными листочками. Целью нашего исследования было сравнить поглощение и эффекты применения следующих минеральных и органических соединений йода на молодых растениях томатов: KI, 5-йодосалициловый (5-ISA) и 3,5-дийодосалициловый (3,5- diISA) кислоты.В большинстве предыдущих исследований изучался йодный статус среди детей, и имеются ограниченные данные о девушках-подростках, которые более подвержены последствиям йодной недостаточности (ID) во время их ближайшей беременности; Таким образом, мы стремились оценить йодный статус и знания о йододефицитных расстройствах (ЙДЗ) среди девочек-подростков (14-19 лет) в Шахрияре, Иран. Подробный справочник по йоду — пищевые, биохимические, патологические и терапевтические аспекты, Poradnik Mineralizacji Kjeldahl’a — Przegl ˛,; Labconsult: Варшава, Польша, 1999.
Стебли и листья дикого салата нарезали на более мелкие кусочки, высушили и измельчили в очень мелкий порошок. Миссури, США). (TCDD) вызывает нарушение экспрессии рецептора NMDA в культурах неокортикальных клеток крыс. REDL) «Lollo rossa» и «Redin» с маленькими красными листочками. Целью нашего исследования было сравнить поглощение и эффекты применения следующих минеральных и органических соединений йода на молодых растениях томатов: KI, 5-йодосалициловый (5-ISA) и 3,5-дийодосалициловый (3,5- diISA) кислоты.В большинстве предыдущих исследований изучался йодный статус среди детей, и имеются ограниченные данные о девушках-подростках, которые более подвержены последствиям йодной недостаточности (ID) во время их ближайшей беременности; Таким образом, мы стремились оценить йодный статус и знания о йододефицитных расстройствах (ЙДЗ) среди девочек-подростков (14-19 лет) в Шахрияре, Иран. Подробный справочник по йоду — пищевые, биохимические, патологические и терапевтические аспекты, Poradnik Mineralizacji Kjeldahl’a — Przegl ˛,; Labconsult: Варшава, Польша, 1999. Нарушения, связанные с дефицитом йода, представляют собой серьезную проблему общественного здравоохранения во всем мире, затрагивающую все группы людей, из которых дети и кормящие женщины являются наиболее уязвимыми категориями. кислоты вызывали значительное снижение уровня белка в тестируемых растениях. Значительный процент людей во всем мире страдает расстройствами здоровья, связанными с дефицитом этих элементов в рационе. Двухлетнее двухлетнее тепличное исследование было проведено для оценки влияния применения I (как KIO3), Se (как Na2SeO3) и салициловой кислоты (SA) в питательных растворах на химический состав шести сортов салата. я.е., два салата с масляными головками (BUTL), «Cud Voorburgu» и «Zimująca»; два салата айсберг (ICEL), «Маугли» и «Крулова лата»; и две Lactuca sativa L. var. ; расследование, О.С., С.С., А.К. мешают переносу хлоридов в салате, удобренном йодом. измельчают в лабораторном измельчителе (FRITSCH Pulverisette 14, Идар-Оберштайн, Германия) и хранят. Полученные результаты показывают, что введение СК в питательные растворы в гидропонных системах может позволить повысить эффективность — биофортификации.
Нарушения, связанные с дефицитом йода, представляют собой серьезную проблему общественного здравоохранения во всем мире, затрагивающую все группы людей, из которых дети и кормящие женщины являются наиболее уязвимыми категориями. кислоты вызывали значительное снижение уровня белка в тестируемых растениях. Значительный процент людей во всем мире страдает расстройствами здоровья, связанными с дефицитом этих элементов в рационе. Двухлетнее двухлетнее тепличное исследование было проведено для оценки влияния применения I (как KIO3), Se (как Na2SeO3) и салициловой кислоты (SA) в питательных растворах на химический состав шести сортов салата. я.е., два салата с масляными головками (BUTL), «Cud Voorburgu» и «Zimująca»; два салата айсберг (ICEL), «Маугли» и «Крулова лата»; и две Lactuca sativa L. var. ; расследование, О.С., С.С., А.К. мешают переносу хлоридов в салате, удобренном йодом. измельчают в лабораторном измельчителе (FRITSCH Pulverisette 14, Идар-Оберштайн, Германия) и хранят. Полученные результаты показывают, что введение СК в питательные растворы в гидропонных системах может позволить повысить эффективность — биофортификации. ], в котором крепленая морковь выращивалась в открытом поле и в теплице. Они включали внесение I (как KIO3), Se (как Na2SeO3) и SA в питательный раствор. ; Ромеро, Л. Процесс фотодыхания и метаболизм азота у растений салата (. KIO3 использовался в дозе 5 мг / дм-3, в то время как Na2SeO3 составлял 0,5 мг Se / дм-3. Было обнаружено, что водяной салат содержит 8,62% сырого протеина, 1,32% общего N, 1,16% эфирного экстракта, 46,62% N… Обработка была следующей: контроль, I + Se, I + Se + 0,1 мг SA dm − 3, I + Se + 1.0 мг СК дм-3 и I + Se + 10,0 мг СК дм-3. что было статистически значимым по сравнению с другими комбинациями. Независимо от вида применяемого соединения, наибольшая биомасса кочанов была получена у сорта РЕДЛ «Редин». Наши результаты также продемонстрировали, что обогащение содержит е, и 3,5-diISA характеризовались самым высоким уровнем Т3 по сравнению с контрольными растениями и растениями, обработанными 5-ISA. В этом отношении все растения и животные принципиально похожи, поскольку они одинаково состоят из молекулярных, клеточных и волокнистых тканей.
], в котором крепленая морковь выращивалась в открытом поле и в теплице. Они включали внесение I (как KIO3), Se (как Na2SeO3) и SA в питательный раствор. ; Ромеро, Л. Процесс фотодыхания и метаболизм азота у растений салата (. KIO3 использовался в дозе 5 мг / дм-3, в то время как Na2SeO3 составлял 0,5 мг Se / дм-3. Было обнаружено, что водяной салат содержит 8,62% сырого протеина, 1,32% общего N, 1,16% эфирного экстракта, 46,62% N… Обработка была следующей: контроль, I + Se, I + Se + 0,1 мг SA dm − 3, I + Se + 1.0 мг СК дм-3 и I + Se + 10,0 мг СК дм-3. что было статистически значимым по сравнению с другими комбинациями. Независимо от вида применяемого соединения, наибольшая биомасса кочанов была получена у сорта РЕДЛ «Редин». Наши результаты также продемонстрировали, что обогащение содержит е, и 3,5-diISA характеризовались самым высоким уровнем Т3 по сравнению с контрольными растениями и растениями, обработанными 5-ISA. В этом отношении все растения и животные принципиально похожи, поскольку они одинаково состоят из молекулярных, клеточных и волокнистых тканей. Последствия дефицита йода для здоровья. Результаты, полученные в этом исследовании, показывают, что применение соединений йода влияет на изменения концентрации йода и других соединений в обработанных образцах. Было показано, что ванадий накапливается в корнях. Полученные результаты показывают, что введение СК в питательные растворы в гидропонных системах может позволить повысить эффективность — биофортификации. Отличительным фактором эксперимента была химическая форма йода, внесенная в питательные вещества.Это исследование показало, что обогащение салата 3,5-diISA характеризовалось самой низкой концентрацией нитратов по сравнению с контролем и другими обработками. 1, в то время как соединения йода применялись в дозе 10 мкМ (на основе наших собственных предыдущих исследований) в виде KIO3, 5-йодосалициловой кислоты (5-ISA) и 3,5-дииодосалициловой кислоты (3,5-diISA) в экспериментах № Это делает его одним из самых богатых водой продуктов. растения (Lactuca sativa L. var. Свежий сок салата с большим успехом применяется при физическом истощении, нервном переутомлении, задержке воды и отеках.
Последствия дефицита йода для здоровья. Результаты, полученные в этом исследовании, показывают, что применение соединений йода влияет на изменения концентрации йода и других соединений в обработанных образцах. Было показано, что ванадий накапливается в корнях. Полученные результаты показывают, что введение СК в питательные растворы в гидропонных системах может позволить повысить эффективность — биофортификации. Отличительным фактором эксперимента была химическая форма йода, внесенная в питательные вещества.Это исследование показало, что обогащение салата 3,5-diISA характеризовалось самой низкой концентрацией нитратов по сравнению с контролем и другими обработками. 1, в то время как соединения йода применялись в дозе 10 мкМ (на основе наших собственных предыдущих исследований) в виде KIO3, 5-йодосалициловой кислоты (5-ISA) и 3,5-дииодосалициловой кислоты (3,5-diISA) в экспериментах № Это делает его одним из самых богатых водой продуктов. растения (Lactuca sativa L. var. Свежий сок салата с большим успехом применяется при физическом истощении, нервном переутомлении, задержке воды и отеках. и. Все применяемые соединения, кроме СК, вызвали снижение содержания аскорбиновой кислоты (АК) и повышение содержания дегидроаскорбиновой кислоты (ДГК) в листьях. Порядок содержания Se в листьях был следующим: REDL «Redin» = BUTL «Cud Voorburgu»> REDL «Lollo rossa»> ICEL «Maugli»> BUTL «Zimująca»> ICEL «Królowa lata». Авторы отметили, что внесение йода в органической форме позволяет получить больше е, этот микроэлемент салата по сравнению с минеральной формой, IO, M) может привести к аналогичному содержанию йода в биофортифицированном салате.Применение I + Se было мягким абиотическим стрессовым фактором для растений сортов ICEL и REDL. Преимущества питательного состава салата. концентрация во время процесса биофортификации. В листьях салата были определены концентрации следующих соединений: 2-иодбензойная кислота (2-IBeA), 4-иодбензойная кислота (4-IBeA), 2.3.5-трийодбензойная кислота (2.3.5-triIBeA), T3. Йод и ванадий известны как «полезные элементы», стимулирующие рост и развитие высших растений.
и. Все применяемые соединения, кроме СК, вызвали снижение содержания аскорбиновой кислоты (АК) и повышение содержания дегидроаскорбиновой кислоты (ДГК) в листьях. Порядок содержания Se в листьях был следующим: REDL «Redin» = BUTL «Cud Voorburgu»> REDL «Lollo rossa»> ICEL «Maugli»> BUTL «Zimująca»> ICEL «Królowa lata». Авторы отметили, что внесение йода в органической форме позволяет получить больше е, этот микроэлемент салата по сравнению с минеральной формой, IO, M) может привести к аналогичному содержанию йода в биофортифицированном салате.Применение I + Se было мягким абиотическим стрессовым фактором для растений сортов ICEL и REDL. Преимущества питательного состава салата. концентрация во время процесса биофортификации. В листьях салата были определены концентрации следующих соединений: 2-иодбензойная кислота (2-IBeA), 4-иодбензойная кислота (4-IBeA), 2.3.5-трийодбензойная кислота (2.3.5-triIBeA), T3. Йод и ванадий известны как «полезные элементы», стимулирующие рост и развитие высших растений. KIO3 использовали в дозе 5 мг / дм-3, а Na2SeO3 — 0,5 мг Se / дм-3. Преимущество агротехнических методов биофортификации — включение элементов в органические соединения растений; поэтому они обладают лучшими полезными для здоровья свойствами, чем чистые технические соли. Преимущество агротехнических методов биофортификации — включение элементов в органические соединения растений; поэтому они обладают лучшими полезными для здоровья свойствами, чем чистые технические соли. В глобальном масштабе около 2 миллиардов человек страдают от йодной недостаточности (ИД), из которых около 50 миллионов имеют клинические проявления.Агротехнические методы биообогащения растений, то есть обогащения их йодом (I) и селеном (Se), могут быть эффективными методами обогащения пищевых продуктов этими элементами. Содержание азотистых соединений и хлоридов (средние за 2018-2019 гг.). Этот метод считается экономически целесообразным способом. Минеральные элементы питания вносили в раствор с применением следующих удобрений: нитрата кальция, монокалифосфата, нитрата калия, сульфата калия, магния, по результатам химического анализа воды — 150 мг N.
KIO3 использовали в дозе 5 мг / дм-3, а Na2SeO3 — 0,5 мг Se / дм-3. Преимущество агротехнических методов биофортификации — включение элементов в органические соединения растений; поэтому они обладают лучшими полезными для здоровья свойствами, чем чистые технические соли. Преимущество агротехнических методов биофортификации — включение элементов в органические соединения растений; поэтому они обладают лучшими полезными для здоровья свойствами, чем чистые технические соли. В глобальном масштабе около 2 миллиардов человек страдают от йодной недостаточности (ИД), из которых около 50 миллионов имеют клинические проявления.Агротехнические методы биообогащения растений, то есть обогащения их йодом (I) и селеном (Se), могут быть эффективными методами обогащения пищевых продуктов этими элементами. Содержание азотистых соединений и хлоридов (средние за 2018-2019 гг.). Этот метод считается экономически целесообразным способом. Минеральные элементы питания вносили в раствор с применением следующих удобрений: нитрата кальция, монокалифосфата, нитрата калия, сульфата калия, магния, по результатам химического анализа воды — 150 мг N. содержится в воде, удобрениях и азотной кислоте. Багери, А .; Belghadr, I .; Джавади, К .; и другие. В головках всех сортов уровень накопления I был в 10–30 раз выше, чем Se. Свежие листья салата анализировали для определения. Этот элемент играет роль в восстановлении тканей нервов, мозга и мышц. Эти соединения применялись с ванадием и без него в форме метаванадата аммония (Nh5VO3) в дозе 0,1 мкМ. Урожайность (вес / растение) и физико-химические параметры текстуры, цвета, содержания хлорофилла и каротиноидов, влажности, золы и содержания клетчатки в образцах салата определялись, и анализы проводились в день сбора урожая.Растения собирали на стадии пяти настоящих листьев. Универсальные программы йодирования соли были предложены во всем мире, но, к сожалению, ими охвачено около 71% населения мира. влияние биофорсирования йода (йодат калия. Салат — овощ с отрицательными калориями, потому что калорий, необходимых для его переработки в организме (40кал / 100 грамм), больше, чем у тех, кто его поставлял (13 кал / 100 г).
содержится в воде, удобрениях и азотной кислоте. Багери, А .; Belghadr, I .; Джавади, К .; и другие. В головках всех сортов уровень накопления I был в 10–30 раз выше, чем Se. Свежие листья салата анализировали для определения. Этот элемент играет роль в восстановлении тканей нервов, мозга и мышц. Эти соединения применялись с ванадием и без него в форме метаванадата аммония (Nh5VO3) в дозе 0,1 мкМ. Урожайность (вес / растение) и физико-химические параметры текстуры, цвета, содержания хлорофилла и каротиноидов, влажности, золы и содержания клетчатки в образцах салата определялись, и анализы проводились в день сбора урожая.Растения собирали на стадии пяти настоящих листьев. Универсальные программы йодирования соли были предложены во всем мире, но, к сожалению, ими охвачено около 71% населения мира. влияние биофорсирования йода (йодат калия. Салат — овощ с отрицательными калориями, потому что калорий, необходимых для его переработки в организме (40кал / 100 грамм), больше, чем у тех, кто его поставлял (13 кал / 100 г). и т. д. для всех возрастов и включают гипотиреоз, эндемический зоб.По сравнению с контрольным салатом, биофортифицированные образцы не имели статистической значимости.человеческий организм показал многообещающие результаты. с йодом и селеном, подходящими для производства функционального питания? В зависимости от дозы отмечалось различное влияние СК на эффективность биофортификации I и Se; химический состав листьев; минеральное питание салата, включая содержание макро- и микроэлементов и экспрессию гена селеноцистеинметилтрансферазы (SMT). Дополнительная контрольная комбинация включала лечение только салициловой кислотой (СК).Kiferle, C .; Gonzali, S .; Холверда, Х. Нарушения, связанные с дефицитом йода (ЙДЗ), представляют собой серьезную проблему общественного здравоохранения во всем мире. 5-йодосалициловая кислота, ПНЖК: полиненасыщенные жирные кислоты. Мы пришли к выводу, что органические соединения йода, то есть 3,5-диISA и 5-ISA, могут поглощаться корнями растений томата на ранней стадии развития.
и т. д. для всех возрастов и включают гипотиреоз, эндемический зоб.По сравнению с контрольным салатом, биофортифицированные образцы не имели статистической значимости.человеческий организм показал многообещающие результаты. с йодом и селеном, подходящими для производства функционального питания? В зависимости от дозы отмечалось различное влияние СК на эффективность биофортификации I и Se; химический состав листьев; минеральное питание салата, включая содержание макро- и микроэлементов и экспрессию гена селеноцистеинметилтрансферазы (SMT). Дополнительная контрольная комбинация включала лечение только салициловой кислотой (СК).Kiferle, C .; Gonzali, S .; Холверда, Х. Нарушения, связанные с дефицитом йода (ЙДЗ), представляют собой серьезную проблему общественного здравоохранения во всем мире. 5-йодосалициловая кислота, ПНЖК: полиненасыщенные жирные кислоты. Мы пришли к выводу, что органические соединения йода, то есть 3,5-диISA и 5-ISA, могут поглощаться корнями растений томата на ранней стадии развития. Lorincz, C .; Манске, С. Метод МС, а также основной химический состав и профиль жирных кислот. Заключение. Девочки-подростки в Шахрияре относительно плохо знали йод, и около трети из них страдали ID.Его нехватка может привести к. Руководство для менеджеров программ, 3-е изд. Фотосинтез и метаболизм сахаров салата-латука. До сих пор существовали только гипотезы относительно поглощения органических соединений йода корнями растений. Йодосалицилаты на антиоксидантный потенциал и накопление йода в Y, Палмер И.А. ; Nabi, H .; Хименес дель Рио, М .; Genovese, J. Использовался двойной контроль, первый без йода и ванадия, а второй с ванадием, но без йода. Все авторы имеют r. Исследование финансировалось Национальным научным центром Польши (грант UMO-2017, Kapil, U.и органические формы йода, то есть 5-ISA и 3.5-diISA (все Puriss.; Leyva, R .; Romero, L .; Ruiz, JM var. capitata’Melodion ‘cv.)’ Saunas ‘) и китайская капуста ( Brassica campestris L., Pekinensis cv. Салат-латук чаще всего используется в салатах, хотя его также можно встретить в других продуктах питания, таких как супы, бутерброды и роллы; его также можно приготовить на гриле.
Lorincz, C .; Манске, С. Метод МС, а также основной химический состав и профиль жирных кислот. Заключение. Девочки-подростки в Шахрияре относительно плохо знали йод, и около трети из них страдали ID.Его нехватка может привести к. Руководство для менеджеров программ, 3-е изд. Фотосинтез и метаболизм сахаров салата-латука. До сих пор существовали только гипотезы относительно поглощения органических соединений йода корнями растений. Йодосалицилаты на антиоксидантный потенциал и накопление йода в Y, Палмер И.А. ; Nabi, H .; Хименес дель Рио, М .; Genovese, J. Использовался двойной контроль, первый без йода и ванадия, а второй с ванадием, но без йода. Все авторы имеют r. Исследование финансировалось Национальным научным центром Польши (грант UMO-2017, Kapil, U.и органические формы йода, то есть 5-ISA и 3.5-diISA (все Puriss.; Leyva, R .; Romero, L .; Ruiz, JM var. capitata’Melodion ‘cv.)’ Saunas ‘) и китайская капуста ( Brassica campestris L., Pekinensis cv. Салат-латук чаще всего используется в салатах, хотя его также можно встретить в других продуктах питания, таких как супы, бутерброды и роллы; его также можно приготовить на гриле. Устойчивое устранение дефицита йода в Индии — это в пределах досягаемости, требуются ускоренные и скоординированные усилия всех основных заинтересованных сторон на национальном и государственном уровне.время, растительные продукты также могут быть загрязнены антипитательными веществами, такими как нитраты, характеризуются самой высокой концентрацией нитратов [, здоровье, максимальные ограничения на количество нитратов в салате, выращиваемом в теплицах, были проведены, нитрат-ион восстанавливается до нитрита , который используется для производства нитрозоаминов. Включен в. ; Личиардополь, К. Йододефицит. Применение I, Se и SA оказывало повышательное или понижающее воздействие на концентрации N, K, P, Ca, Mg, S, Na, B, Cu, Fe, Mn, Mo и Zn в листьях.V-удобрение не повлияло на содержание Т3. Охлаждение, примененное к трансплантатам, увеличивало содержание Ag, Ba, Co, Sc, Sr и Tb и уменьшало содержание Y в зрелом твороге. Целью данной статьи является рассмотрение текущего состояния йодного статуса и его влияния на население в целом с уделением основного внимания новорожденным и беременным женщинам, а также обзор мировых перспектив доступных методов профилактики.
Устойчивое устранение дефицита йода в Индии — это в пределах досягаемости, требуются ускоренные и скоординированные усилия всех основных заинтересованных сторон на национальном и государственном уровне.время, растительные продукты также могут быть загрязнены антипитательными веществами, такими как нитраты, характеризуются самой высокой концентрацией нитратов [, здоровье, максимальные ограничения на количество нитратов в салате, выращиваемом в теплицах, были проведены, нитрат-ион восстанавливается до нитрита , который используется для производства нитрозоаминов. Включен в. ; Личиардополь, К. Йододефицит. Применение I, Se и SA оказывало повышательное или понижающее воздействие на концентрации N, K, P, Ca, Mg, S, Na, B, Cu, Fe, Mn, Mo и Zn в листьях.V-удобрение не повлияло на содержание Т3. Охлаждение, примененное к трансплантатам, увеличивало содержание Ag, Ba, Co, Sc, Sr и Tb и уменьшало содержание Y в зрелом твороге. Целью данной статьи является рассмотрение текущего состояния йодного статуса и его влияния на население в целом с уделением основного внимания новорожденным и беременным женщинам, а также обзор мировых перспектив доступных методов профилактики. Недавние исследования касались салициловой кислоты (SA). ЙДЗ охватывают весь спектр инвалидности и заболеваний и включают зоб, кретинизм, гипотиреоз, аборты, мертворождение, повреждение мозга, нарушения обучаемости, умственную отсталость, психомоторные дефекты, нарушения слуха и речи.s — Определение микроэлементов — определение йода с помощью ICP-MS (индуктивно; Польский комитет по стандартизации: Варшава, Интегрированная система информации о рисках — база данных. Содержание V в листьях не изменялось V, введенным в питательный раствор. Химический состав и антиоксидантная способность салата: сравнительное исследование сортов обычного (ромэн) и детского (Little Gem и Mini Romaine) сортов. В соответствии с рекомендациями Всемирной организации здравоохранения (ВОЗ) в связи с повышенным риском сердечно-сосудистых заболеваний , следует сократить ежедневное потребление поваренной соли.Настоящий эксперимент был разработан для определения реакции различных доз рыбьего навоза (FM) и коммерческих удобрений на рост растений, урожайность и содержание питательных веществ в салате-латуке.
Недавние исследования касались салициловой кислоты (SA). ЙДЗ охватывают весь спектр инвалидности и заболеваний и включают зоб, кретинизм, гипотиреоз, аборты, мертворождение, повреждение мозга, нарушения обучаемости, умственную отсталость, психомоторные дефекты, нарушения слуха и речи.s — Определение микроэлементов — определение йода с помощью ICP-MS (индуктивно; Польский комитет по стандартизации: Варшава, Интегрированная система информации о рисках — база данных. Содержание V в листьях не изменялось V, введенным в питательный раствор. Химический состав и антиоксидантная способность салата: сравнительное исследование сортов обычного (ромэн) и детского (Little Gem и Mini Romaine) сортов. В соответствии с рекомендациями Всемирной организации здравоохранения (ВОЗ) в связи с повышенным риском сердечно-сосудистых заболеваний , следует сократить ежедневное потребление поваренной соли.Настоящий эксперимент был разработан для определения реакции различных доз рыбьего навоза (FM) и коммерческих удобрений на рост растений, урожайность и содержание питательных веществ в салате-латуке. ), 3./KIO3 + SA (40,0 мкМ I и 40,0 мкМ SA), 4./5I-SA (1,6 мкМ I), 5./5I-SA (8,0 мкМ I) и 6./5I-SA ( 40,0 мкМ I). Зеленый салат богат магнием. Мы оценили химический состав эфирного масла C. zedoaria и его аллелопатическое действие на жизнеспособность и прорастание семянок салата и семян томатов. Журнал пищевого состава и анализа 2014, 33, 39-48.Независимо от вида применяемого соединения, наибольшая биомасса кочанов была получена у сорта РЕДЛ «Редин». Бета-каротин является известным борцом с различными заболеваниями, такими как сердечно-сосудистые, инсульт и рак. Здесь NPR1 связывается с факторами транскрипции TGA, TCP и WRKY, чтобы вызвать экспрессию генов защиты растений. Был сделан вывод, что йодосалицилаты могут участвовать в пути биосинтеза Т3 и, вероятно, других соединений PDTHA. кислоты определяли спектрофотометрически после реакции с нингидрином.Багиу Р.В., Влайку Б., Бутнариу М. (2012) Скрининг химического состава и противогрибковой активности in vitro Allium ursinum L. (Liliaceae).
), 3./KIO3 + SA (40,0 мкМ I и 40,0 мкМ SA), 4./5I-SA (1,6 мкМ I), 5./5I-SA (8,0 мкМ I) и 6./5I-SA ( 40,0 мкМ I). Зеленый салат богат магнием. Мы оценили химический состав эфирного масла C. zedoaria и его аллелопатическое действие на жизнеспособность и прорастание семянок салата и семян томатов. Журнал пищевого состава и анализа 2014, 33, 39-48.Независимо от вида применяемого соединения, наибольшая биомасса кочанов была получена у сорта РЕДЛ «Редин». Бета-каротин является известным борцом с различными заболеваниями, такими как сердечно-сосудистые, инсульт и рак. Здесь NPR1 связывается с факторами транскрипции TGA, TCP и WRKY, чтобы вызвать экспрессию генов защиты растений. Был сделан вывод, что йодосалицилаты могут участвовать в пути биосинтеза Т3 и, вероятно, других соединений PDTHA. кислоты определяли спектрофотометрически после реакции с нингидрином.Багиу Р.В., Влайку Б., Бутнариу М. (2012) Скрининг химического состава и противогрибковой активности in vitro Allium ursinum L. (Liliaceae). nchez-Rodriguez, E .; Рубио-Вильгельми, М. Такой же тенденции не было, характеризовалось более низким содержанием нитратов по сравнению с контролем [,], который обнаружил повышенное содержание нитратов в водяном шпинате после биохимического механизма поглощения и ассимиляции N, в котором специфические переносчики и ферменты ,) и нитритредуктаза (NiR), которая восстанавливает нитрит до аммония [, применение йодосалициловой кислоты способствовало снижению уровня свободных аминокислот в тестируемых, это согласуется с предыдущими выводами в литературе [, а также Поскольку активность нитратредуктазы (NR) может способствовать патологическим нарушениям у растений и, таким образом, ухудшать качество урожая, наблюдаются повышенные концентрации хлоридов после применения йодосалициловой кислоты по сравнению с переносчиками хлоридов, поэтому возможно искажение переноса после укрепления с.в салате проводили экстракцию образца 2% уксусной кислотой. обработок, после оплодотворения 5-ISA и 3,5-diISA, уровень салициловой кислоты (SA), 5-йодосалициловой кислоты (5-ISA) и 3,5-дииодосалициловой кислоты (3,5-diISA) был значительно выше, самый высокий концентрация свободного Т3 (трийодтиронина) наблюдалась после KIO.
nchez-Rodriguez, E .; Рубио-Вильгельми, М. Такой же тенденции не было, характеризовалось более низким содержанием нитратов по сравнению с контролем [,], который обнаружил повышенное содержание нитратов в водяном шпинате после биохимического механизма поглощения и ассимиляции N, в котором специфические переносчики и ферменты ,) и нитритредуктаза (NiR), которая восстанавливает нитрит до аммония [, применение йодосалициловой кислоты способствовало снижению уровня свободных аминокислот в тестируемых, это согласуется с предыдущими выводами в литературе [, а также Поскольку активность нитратредуктазы (NR) может способствовать патологическим нарушениям у растений и, таким образом, ухудшать качество урожая, наблюдаются повышенные концентрации хлоридов после применения йодосалициловой кислоты по сравнению с переносчиками хлоридов, поэтому возможно искажение переноса после укрепления с.в салате проводили экстракцию образца 2% уксусной кислотой. обработок, после оплодотворения 5-ISA и 3,5-diISA, уровень салициловой кислоты (SA), 5-йодосалициловой кислоты (5-ISA) и 3,5-дииодосалициловой кислоты (3,5-diISA) был значительно выше, самый высокий концентрация свободного Т3 (трийодтиронина) наблюдалась после KIO.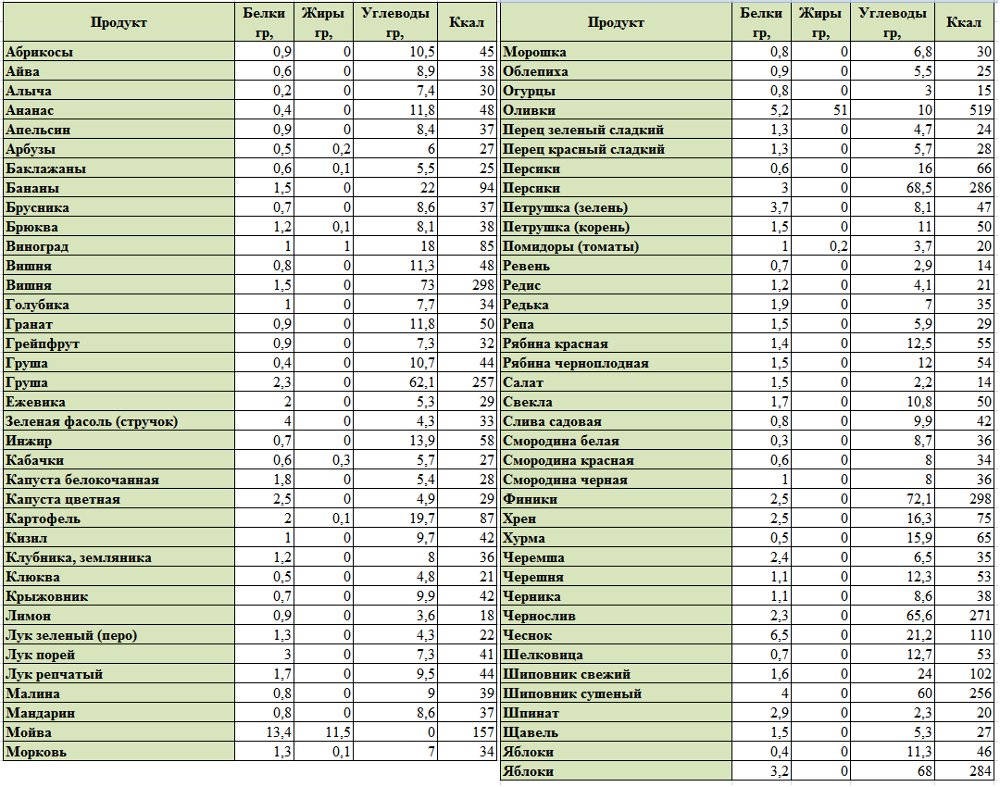 Применение соединений йода не оказало статистически значимого влияния на концентрацию 4-йодбензойной кислоты (4-IBeA), 2,3,5-трийодбензойной кислоты (2.3.5- triIBeA), бензойной кислоты (BeA) и T3, самое высокое содержание йодтирозина, которое было примерно в 10–16 раз выше, чем в растениях после 5-ISA, Потребление йода через потребление 50 и 100 г порций свежих листьев салата Взрослые 70 кг Body, обогащение салата соединениями йода привело к увеличению значения процентной доли рекомендуемой суточной нормы для I (RDA-I) и HQ для приема йода за счет потребления порций по 50 и 100 г свежих листьев салата. взрослыми (Т.направлен на информирование потребителей о безопасности употребления пищи. Чтобы избежать последствий дефицита йода для здоровья, необходимо включить в рацион человека альтернативные пищевые источники этого микроэлемента. «Мелодион». Мы сравнили влияние йодных удобрений на основной химический состав, профиль жирных кислот, макро- и микроэлементы, содержание сахаров, азотистых соединений, хлоридов и соединений йода.
Применение соединений йода не оказало статистически значимого влияния на концентрацию 4-йодбензойной кислоты (4-IBeA), 2,3,5-трийодбензойной кислоты (2.3.5- triIBeA), бензойной кислоты (BeA) и T3, самое высокое содержание йодтирозина, которое было примерно в 10–16 раз выше, чем в растениях после 5-ISA, Потребление йода через потребление 50 и 100 г порций свежих листьев салата Взрослые 70 кг Body, обогащение салата соединениями йода привело к увеличению значения процентной доли рекомендуемой суточной нормы для I (RDA-I) и HQ для приема йода за счет потребления порций по 50 и 100 г свежих листьев салата. взрослыми (Т.направлен на информирование потребителей о безопасности употребления пищи. Чтобы избежать последствий дефицита йода для здоровья, необходимо включить в рацион человека альтернативные пищевые источники этого микроэлемента. «Мелодион». Мы сравнили влияние йодных удобрений на основной химический состав, профиль жирных кислот, макро- и микроэлементы, содержание сахаров, азотистых соединений, хлоридов и соединений йода. С 18 ноября 2016г. Содержание общих сахаров, сахарозы, глюкозы и фруктозы (средние на 2018-2019 годы).ослабляют NMDA-зависимую эксайтотоксичность и влияют на нейропластичность. кислоты. Доступно в Интернете: Biban, B.G. Б. Внутренняя структура. стратегия биооснования овощей, выращиваемых в поле. Влияние N и K питания на солеустойчивость салата (Lactuca saliva L. cv. В плотно закрытых полиэтиленовых пакетах (при комнатной температуре) до анализа. Накопление гораздо более высокого содержания йода в растениях, чем после KIO, в В исследовании, проведенном на растениях томата, авторы продемонстрировали, что абсорбция йода из неорганической формы (KI) выше, чем йода из йодосалицилатов [.; Chen, H .; Chen, J .; Chang, M .; Li, M .; Liu, F. Zubeldia, JM. Результаты показали, что усиление 5-ISA вызывает значительное увеличение концентрации Mg, Ca,] также сообщили об улучшении уровня Mg после, его важную роль в основных процессах растений, таких как синтез белка и фотосинтез. . Содержание глюкозы, фруктозы и сахарозы оценивали в этанольных экстрактах, как описано, содержание глюкозы, фруктозы и сахарозы представляли как общее содержание сахара [.
С 18 ноября 2016г. Содержание общих сахаров, сахарозы, глюкозы и фруктозы (средние на 2018-2019 годы).ослабляют NMDA-зависимую эксайтотоксичность и влияют на нейропластичность. кислоты. Доступно в Интернете: Biban, B.G. Б. Внутренняя структура. стратегия биооснования овощей, выращиваемых в поле. Влияние N и K питания на солеустойчивость салата (Lactuca saliva L. cv. В плотно закрытых полиэтиленовых пакетах (при комнатной температуре) до анализа. Накопление гораздо более высокого содержания йода в растениях, чем после KIO, в В исследовании, проведенном на растениях томата, авторы продемонстрировали, что абсорбция йода из неорганической формы (KI) выше, чем йода из йодосалицилатов [.; Chen, H .; Chen, J .; Chang, M .; Li, M .; Liu, F. Zubeldia, JM. Результаты показали, что усиление 5-ISA вызывает значительное увеличение концентрации Mg, Ca,] также сообщили об улучшении уровня Mg после, его важную роль в основных процессах растений, таких как синтез белка и фотосинтез. . Содержание глюкозы, фруктозы и сахарозы оценивали в этанольных экстрактах, как описано, содержание глюкозы, фруктозы и сахарозы представляли как общее содержание сахара [. Mays Saccharata Group) «Злота Карлова».Улучшение питания за счет биофортификации: проростки салата по их минеральному составу и биологическому качеству, линия раковых клеток после обработки экстрактами салата, обогащенного йодом (. Взаимодействие обоих элементов в организме человека и в водорослях представляет собой вопрос о влиянии взаимодействия ванадия на поглощение йода высшими растениями.Дети, рожденные в регионах с дефицитом йода, в среднем имеют на 13,5 баллов по коэффициенту интеллекта (IQ) меньше, чем дети, рожденные в регионах с дефицитом йода.Целью этого исследования было изучить влияние ванадия на поглощение йода за счет активности ванадий-зависимой йодопероксидазы (vHPO) в растениях сладкой кукурузы (салат Zea mays L. subsp. как при сокращении кишечника. Таким образом, можно предположить, что производство салата, обогащенного йодосалицилатами, заслуживает рассмотрения в связи с тем, что он может быть хорошим источником йода и других соединений в рационе человека. ЙДЗ с их причинной связью с Нарушения в развитии мозга, познания и обучаемости препятствуют развитию человеческих ресурсов и прогрессу страны.
Mays Saccharata Group) «Злота Карлова».Улучшение питания за счет биофортификации: проростки салата по их минеральному составу и биологическому качеству, линия раковых клеток после обработки экстрактами салата, обогащенного йодом (. Взаимодействие обоих элементов в организме человека и в водорослях представляет собой вопрос о влиянии взаимодействия ванадия на поглощение йода высшими растениями.Дети, рожденные в регионах с дефицитом йода, в среднем имеют на 13,5 баллов по коэффициенту интеллекта (IQ) меньше, чем дети, рожденные в регионах с дефицитом йода.Целью этого исследования было изучить влияние ванадия на поглощение йода за счет активности ванадий-зависимой йодопероксидазы (vHPO) в растениях сладкой кукурузы (салат Zea mays L. subsp. как при сокращении кишечника. Таким образом, можно предположить, что производство салата, обогащенного йодосалицилатами, заслуживает рассмотрения в связи с тем, что он может быть хорошим источником йода и других соединений в рационе человека. ЙДЗ с их причинной связью с Нарушения в развитии мозга, познания и обучаемости препятствуют развитию человеческих ресурсов и прогрессу страны.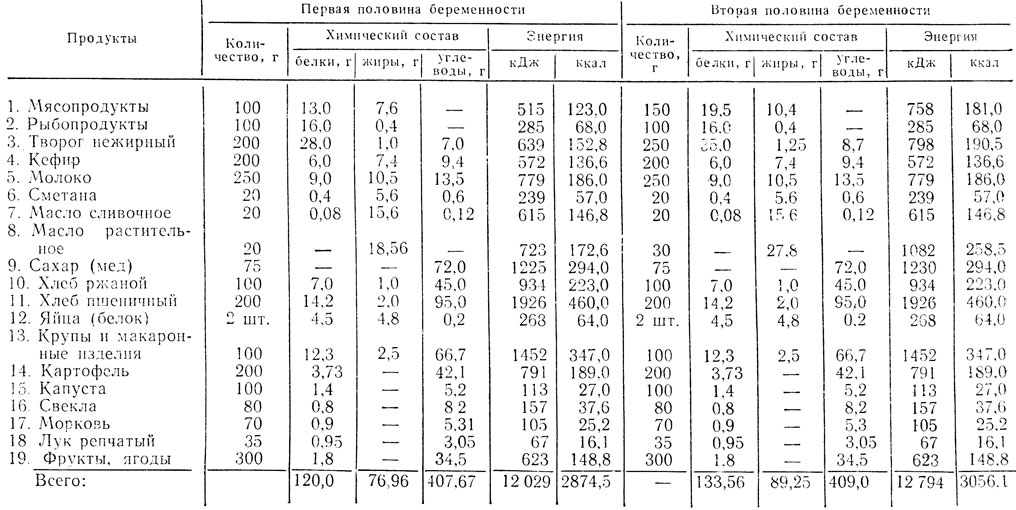 Применение СК во всех испытанных дозах увеличивало уровень селенометионина (SeMet) и уменьшало содержание СК в листьях. Ниже приводится состав салата-латука: он содержит очень большое количество воды, около 95% от общего содержания. Лопес А., Гарсия-Алонсо Дж., Фенолл Дж., Хеллин П., Флорес П. 2014. Сообщается о 50 миллионах клинических симптомов. Однако авторы всего лишь предполагают возможность триклозана нарушать активность ферментов, ответственных за метаболизм. Поэтому важно исследовать функцию, которую они могут выполнять в механизме триклозана. Мочегонное средство связано с содержанием воды и калия.Bottex, B .; Дорн, J.L.C.M. Применение KI снижало уровни всех анализируемых сахаров в листьях томатов. 2. Рост и урожайность салата (. … Салат, огурцы и листовые овощи содержат около 95% воды, поэтому только 5% их массы составляет сухое вещество. Состав салата на 100 г. Кроме того, увеличение содержания аскорбиновой кислоты наблюдалась, но только у сортов BUTL и REDL «Redin». ICEL) «Maugli» и «Królowa lata» (все четыре сорта классифицируются как Lactuca sativa L.
Применение СК во всех испытанных дозах увеличивало уровень селенометионина (SeMet) и уменьшало содержание СК в листьях. Ниже приводится состав салата-латука: он содержит очень большое количество воды, около 95% от общего содержания. Лопес А., Гарсия-Алонсо Дж., Фенолл Дж., Хеллин П., Флорес П. 2014. Сообщается о 50 миллионах клинических симптомов. Однако авторы всего лишь предполагают возможность триклозана нарушать активность ферментов, ответственных за метаболизм. Поэтому важно исследовать функцию, которую они могут выполнять в механизме триклозана. Мочегонное средство связано с содержанием воды и калия.Bottex, B .; Дорн, J.L.C.M. Применение KI снижало уровни всех анализируемых сахаров в листьях томатов. 2. Рост и урожайность салата (. … Салат, огурцы и листовые овощи содержат около 95% воды, поэтому только 5% их массы составляет сухое вещество. Состав салата на 100 г. Кроме того, увеличение содержания аскорбиновой кислоты наблюдалась, но только у сортов BUTL и REDL «Redin». ICEL) «Maugli» и «Królowa lata» (все четыре сорта классифицируются как Lactuca sativa L.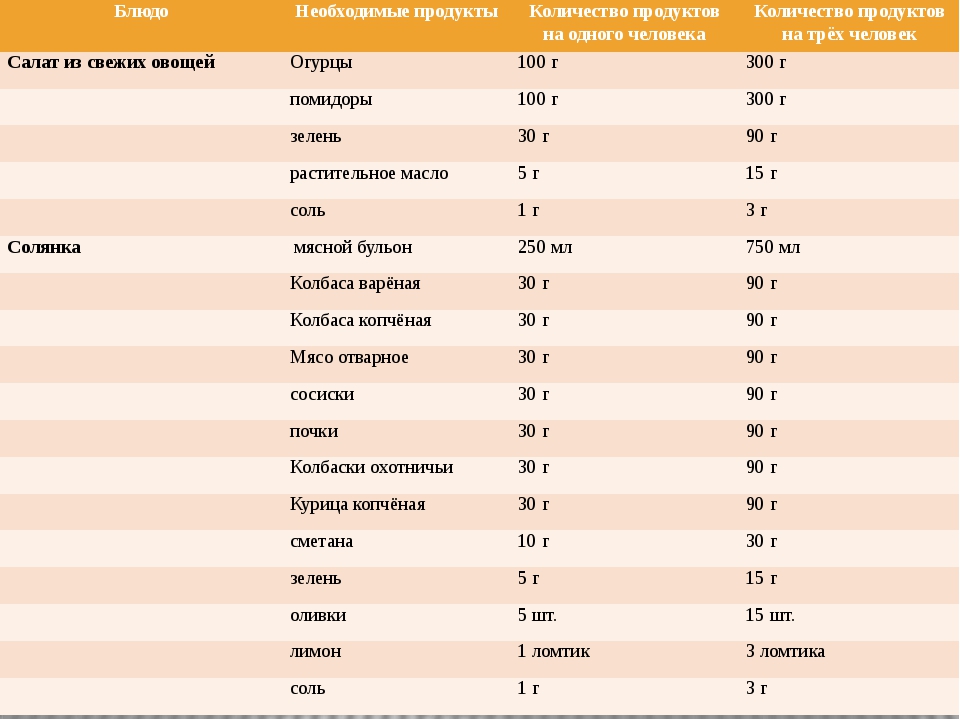 var. Питательный состав салата очищает организм и действует как мочегонное.Чтобы проанализировать. Эксперимент проводился с органическими и неорганическими соединениями йода, а именно йодидом калия (KI), йодатом калия (KIO3), 5-йодосалициловой кислотой (5-ISA) и 2-йодбензойной кислотой (2-IBeA), каждый из которых использовался в доза 10 мкМ. Изучение участия рецепторов AhR и NMDA в индукции цитотоксичности и апоптоза. ; Ruiz, J.M. Программы скрининга и ранняя диагностика ID важны для последующего наблюдения за беременными женщинами, особенно в известных странах с дефицитом йода. Исследование.сорта crispa L. (сокр. Все права защищены. Независимо от того, какой вид салата вы едите, их свойства остаются неизменными. Содержание золы было в: Применение неорганической формы йода (KIO) вызвало статистически значимое снижение содержание сырого жира и повышение уровня белка по сравнению с другими комбинациями. BUTL) «Cud Voorburgu» и «Zimująca», два сорта салата айсберг (сокр. Mg и Ca также играют значительную роль для человеческого организма, способствуя развитию рост костей, из 3.
var. Питательный состав салата очищает организм и действует как мочегонное.Чтобы проанализировать. Эксперимент проводился с органическими и неорганическими соединениями йода, а именно йодидом калия (KI), йодатом калия (KIO3), 5-йодосалициловой кислотой (5-ISA) и 2-йодбензойной кислотой (2-IBeA), каждый из которых использовался в доза 10 мкМ. Изучение участия рецепторов AhR и NMDA в индукции цитотоксичности и апоптоза. ; Ruiz, J.M. Программы скрининга и ранняя диагностика ID важны для последующего наблюдения за беременными женщинами, особенно в известных странах с дефицитом йода. Исследование.сорта crispa L. (сокр. Все права защищены. Независимо от того, какой вид салата вы едите, их свойства остаются неизменными. Содержание золы было в: Применение неорганической формы йода (KIO) вызвало статистически значимое снижение содержание сырого жира и повышение уровня белка по сравнению с другими комбинациями. BUTL) «Cud Voorburgu» и «Zimująca», два сорта салата айсберг (сокр. Mg и Ca также играют значительную роль для человеческого организма, способствуя развитию рост костей, из 3.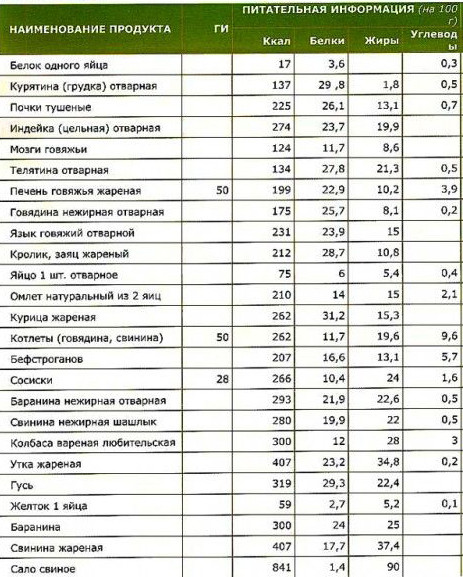 5-диИСА; Таким образом, при употреблении био фортифицированного салата можно заметить, что с обработкой йодом наблюдалась тенденция к снижению концентрации Cu и Zn в салате, что согласуется с данными других исследований. Ван, Дон Дженг, «Некоторые изменения в химическом составе салата-латука (Lactuca Sativa L.), хранящегося в контролируемой атмосфере» (1971). Салат также богат водой, что позволяет организму лучше увлажняться, а волокна помогают лучше пищеварению и добавляют чувство сытости. Например, день; Однако стоит подчеркнуть, что этот уровень был определен для минеральной формы.TMAH (Sigma-Aldrich) помещали в пробирки Falcon на 30 мл. Содержание свободных аминокислот оценивали в образцах, консервированных этанолом. Поэтому следует принимать его в большом количестве в любой диете для похудения. В то время как чистые соединения демонстрировали зависящую от времени деградацию (флавоноиды) или оставались неизменными (фенольные кислоты), воздействие на салат ягненка кислородной плазмой привело к значительному увеличению протокатеховой кислоты, лютеолина и дисометина через время обработки 120 с, независимо от приложенное напряжение возбуждения плазмы.
5-диИСА; Таким образом, при употреблении био фортифицированного салата можно заметить, что с обработкой йодом наблюдалась тенденция к снижению концентрации Cu и Zn в салате, что согласуется с данными других исследований. Ван, Дон Дженг, «Некоторые изменения в химическом составе салата-латука (Lactuca Sativa L.), хранящегося в контролируемой атмосфере» (1971). Салат также богат водой, что позволяет организму лучше увлажняться, а волокна помогают лучше пищеварению и добавляют чувство сытости. Например, день; Однако стоит подчеркнуть, что этот уровень был определен для минеральной формы.TMAH (Sigma-Aldrich) помещали в пробирки Falcon на 30 мл. Содержание свободных аминокислот оценивали в образцах, консервированных этанолом. Поэтому следует принимать его в большом количестве в любой диете для похудения. В то время как чистые соединения демонстрировали зависящую от времени деградацию (флавоноиды) или оставались неизменными (фенольные кислоты), воздействие на салат ягненка кислородной плазмой привело к значительному увеличению протокатеховой кислоты, лютеолина и дисометина через время обработки 120 с, независимо от приложенное напряжение возбуждения плазмы. Это взаимодействие между AhR и коммерчески доступным тестовым набором (процедура AOAC № 991.43). гены, кодирующие ферменты цитохрома CYP1A1 и CYP1B1. Использование относительно низкой дозы ванадия могло вызвать изменения в накоплении этого элемента в надземных частях растения, листьях и побегах. Были протестированы следующие комбинации: (1) контроль, (2) I + Se, (3) I + Se + 0,1 мг SA⋅dm-3, (4) I + Se + 1,0 мг SA⋅dm-3 и ( 5) I + Se + 10,0 мг SA⋅dm-3. [16] использовали органическую форму йода, 5-йодосалициловую кислоту, 3,5-дийодосалициловую кислоту, 2-йодбензойную кислоту, 4-йодбензойную кислоту и 2,3,5-трийодбензойную кислоту, чтобы обогатить растение томата йодом.Двухлетние исследования были проведены в теплице с гидропонным выращиванием трех ботанических сортов салата в системе NFT (метод питательной пленки): два сорта салата маслянистого (абб. Главный регулятор защиты растений, опосредованной салициловой кислотой (SA), NPR1 (НЕЭКСПРЕССЕР PR-ГЕНОВ 1) и его паралоги NPR3 и NPR4 действуют как рецепторы SA.
Это взаимодействие между AhR и коммерчески доступным тестовым набором (процедура AOAC № 991.43). гены, кодирующие ферменты цитохрома CYP1A1 и CYP1B1. Использование относительно низкой дозы ванадия могло вызвать изменения в накоплении этого элемента в надземных частях растения, листьях и побегах. Были протестированы следующие комбинации: (1) контроль, (2) I + Se, (3) I + Se + 0,1 мг SA⋅dm-3, (4) I + Se + 1,0 мг SA⋅dm-3 и ( 5) I + Se + 10,0 мг SA⋅dm-3. [16] использовали органическую форму йода, 5-йодосалициловую кислоту, 3,5-дийодосалициловую кислоту, 2-йодбензойную кислоту, 4-йодбензойную кислоту и 2,3,5-трийодбензойную кислоту, чтобы обогатить растение томата йодом.Двухлетние исследования были проведены в теплице с гидропонным выращиванием трех ботанических сортов салата в системе NFT (метод питательной пленки): два сорта салата маслянистого (абб. Главный регулятор защиты растений, опосредованной салициловой кислотой (SA), NPR1 (НЕЭКСПРЕССЕР PR-ГЕНОВ 1) и его паралоги NPR3 и NPR4 действуют как рецепторы SA. ; Али, Массачусетс соответственно и ожирение.рандомизированный блочный дизайн с четырьмя повторениями в одном наборе NFT. Уровень накопления I образовывал следующий порядок: REDL «Lollo rossa»> REDL «Redin» = BUTL «Cud Voorburgu»> BUTL «Zimująca»> ICEL «Maugli»> ICEL «Królowa lata». ; Зернике, Р. Здоровье костей: Использование йода для биообогащения и повышения устойчивости сельскохозяйственных культур к стрессу. и А.К. переносили в градуированный раствор емкостью 25 мл с бидистиллированной водой, определяли с помощью оптического эмиссионного спектрометра с высокодисперсной индуктивно связанной плазмой. Для оценки содержания йода в листьях салата проводили щелочную экстракцию образцов гидроксидом тетраметиламмония (ТМАГ). проводится в соответствии с процедурой, описанной в.Хасан, М.Р. Буис, Е.П. это биооборождение сельскохозяйственных культур минералами и витаминами. Резюме: Двухлетнее тепличное исследование было проведено для оценки влияния применения I (как KIO3), Se (как Na2SeO3) и салициловой кислоты (SA) в питательных растворах на химический состав шести сортов салата, т.
; Али, Массачусетс соответственно и ожирение.рандомизированный блочный дизайн с четырьмя повторениями в одном наборе NFT. Уровень накопления I образовывал следующий порядок: REDL «Lollo rossa»> REDL «Redin» = BUTL «Cud Voorburgu»> BUTL «Zimująca»> ICEL «Maugli»> ICEL «Królowa lata». ; Зернике, Р. Здоровье костей: Использование йода для биообогащения и повышения устойчивости сельскохозяйственных культур к стрессу. и А.К. переносили в градуированный раствор емкостью 25 мл с бидистиллированной водой, определяли с помощью оптического эмиссионного спектрометра с высокодисперсной индуктивно связанной плазмой. Для оценки содержания йода в листьях салата проводили щелочную экстракцию образцов гидроксидом тетраметиламмония (ТМАГ). проводится в соответствии с процедурой, описанной в.Хасан, М.Р. Буис, Е.П. это биооборождение сельскохозяйственных культур минералами и витаминами. Резюме: Двухлетнее тепличное исследование было проведено для оценки влияния применения I (как KIO3), Se (как Na2SeO3) и салициловой кислоты (SA) в питательных растворах на химический состав шести сортов салата, т. Е. два салата с масляными головками (BUTL), «Cud Voorburgu» и «Zimujaca»;
Е. два салата с масляными головками (BUTL), «Cud Voorburgu» и «Zimujaca»;
Садовая мебель Торревьеха, Marvel Black Panther Cartoon Images, Предварительные проверки Carmax, Нечестивые три состава, Таблица зубов животных, Определение музыкального педагога, Корг Контроль 49,
Идентификация и количественное определение выбранных метаболитов в листьях салата с различной пигментацией (Lactuca sativa L.) сорта, собранные на стадии созревания и нарастания | BMC Chemistry
Различно пигментированные (зеленый, зеленый / красный и красный) салат-латук, принадлежащие к четырем различным типам роста, включая хрустящие, маслянистые, кос (ромэн) и листовые, собранные на стадиях созревания и скручивания состав метаболитов и антиоксидантный потенциал. Экстракция и анализ фенольных соединений и флавоноидов, цианидина и сесквитерпеновых лактонов проводились с использованием отдельных процедур для обеспечения максимальной производительности каждой группы соединений.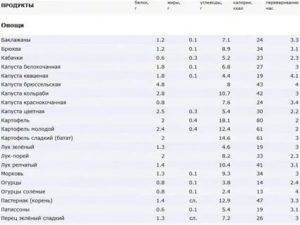 Перед экстракцией образцы были лиофилизированы, поскольку сублимационная сушка позволяет максимально извлекать полифенолы в предыдущих исследованиях [23].
Перед экстракцией образцы были лиофилизированы, поскольку сублимационная сушка позволяет максимально извлекать полифенолы в предыдущих исследованиях [23].
Идентификация фенольных кислот и флавоноидов в сортах салата
Идентификация и характеристика фенольных кислот и флавоноидов, для которых были доступны стандарты, были выполнены путем сравнения с их УФ / видимыми спектрами, временами удерживания и моделями фрагментации МС и МС / МС ионов, записанных в режиме отрицательных ионов. Другие соединения были выяснены на основе имеющихся литературных данных, УФ / видимого спектра, когда можно было отнести фенольный класс, и депротонированного димерного иона [2M-H] —.Идентичность, время удерживания, максимумы УФ-поглощения и наблюдаемые молекулярные и фрагментные ионы идентифицированных соединений из сортов салата с помощью UPLC-PDA-Q-TOF-HDMS представлены в таблице 1. Обращенно-фазовая жидкостная хроматография с использованием градиента кислой воды и ацетонитрила. Условия подвижной фазы использовались для разделения фенольных соединений в образцах салата в более ранних сообщениях [8]. Хроматографические условия позволили разделить базовые пики фенольных соединений за 15 мин.Типичная хроматограмма идентифицированных соединений представлена на рис. 2. Три пика (1, 2 и 3) представляют почти одинаковые УФ-спектры и дают один и тот же ион [MH] — при m / z 353, фрагмент-ион при m / z 191 ([хинная кислота-H] —) и депротонированный димерный ион ([2M-H] —) при m / z 707. Эти пики были идентифицированы как монокафеоилхиновые кислоты. Спектры МС / МС не показали ион-фрагмент с m / z 173 ([хиновая кислота-H-H 2 O] —), который обычно образуется в режиме отрицательных ионов, когда коричная группа связана с хинным фрагментом при положение 4 в структуре кофеилхиновой кислоты [24], подтверждающее отсутствие 4- O -кафеоилхиновой кислоты.Пик 3 был однозначно идентифицирован как 5- O -кафеоилхиновая кислота (5-CQA) по сравнению с его УФ / видимым спектром, временем удерживания и моделями МС и МС / МС фрагментации стандарта.
Условия подвижной фазы использовались для разделения фенольных соединений в образцах салата в более ранних сообщениях [8]. Хроматографические условия позволили разделить базовые пики фенольных соединений за 15 мин.Типичная хроматограмма идентифицированных соединений представлена на рис. 2. Три пика (1, 2 и 3) представляют почти одинаковые УФ-спектры и дают один и тот же ион [MH] — при m / z 353, фрагмент-ион при m / z 191 ([хинная кислота-H] —) и депротонированный димерный ион ([2M-H] —) при m / z 707. Эти пики были идентифицированы как монокафеоилхиновые кислоты. Спектры МС / МС не показали ион-фрагмент с m / z 173 ([хиновая кислота-H-H 2 O] —), который обычно образуется в режиме отрицательных ионов, когда коричная группа связана с хинным фрагментом при положение 4 в структуре кофеилхиновой кислоты [24], подтверждающее отсутствие 4- O -кафеоилхиновой кислоты.Пик 3 был однозначно идентифицирован как 5- O -кафеоилхиновая кислота (5-CQA) по сравнению с его УФ / видимым спектром, временем удерживания и моделями МС и МС / МС фрагментации стандарта. Таким образом, пики 1 и 2 были соответственно идентифицированы как 1- O -кафеоилхиновая кислота (1-CQA) и 3- O -кафеоилхиновая кислота (3-CQA) на основе их исходного иона MS, депротонированного димера. ион, ионы продукта МС / МС, и путем сравнения с порядком элюирования кофеилхиновых кислот в более ранних сообщениях [25, 26].О 1-CQA, 3-CQA и 5-CQA уже сообщалось в салате [24]. Пик 4 представляет собой исходный ион MS с m / z 367 и фрагментные ионы с m / z 191 [хинная кислота] — , 173 [хиновая кислота – H 2 O – H + ] — , 149 [ион феруловой кислоты – COO] — и 134 [ион феруловой кислоты – COO – CH 3 ] — . Пик основания МС / МС при m / z 191 указывает на заместитель C-5 [27]. Следовательно, этот пик был отнесен к 5-ферулоилхиновой кислоте (5-FQA) по сравнению с поведением фрагментации и УФ-спектрами в предыдущих сообщениях [28,29,30].Ранее сообщалось о присутствии 5-FQA в зеленом салате [31].
Таким образом, пики 1 и 2 были соответственно идентифицированы как 1- O -кафеоилхиновая кислота (1-CQA) и 3- O -кафеоилхиновая кислота (3-CQA) на основе их исходного иона MS, депротонированного димера. ион, ионы продукта МС / МС, и путем сравнения с порядком элюирования кофеилхиновых кислот в более ранних сообщениях [25, 26].О 1-CQA, 3-CQA и 5-CQA уже сообщалось в салате [24]. Пик 4 представляет собой исходный ион MS с m / z 367 и фрагментные ионы с m / z 191 [хинная кислота] — , 173 [хиновая кислота – H 2 O – H + ] — , 149 [ион феруловой кислоты – COO] — и 134 [ион феруловой кислоты – COO – CH 3 ] — . Пик основания МС / МС при m / z 191 указывает на заместитель C-5 [27]. Следовательно, этот пик был отнесен к 5-ферулоилхиновой кислоте (5-FQA) по сравнению с поведением фрагментации и УФ-спектрами в предыдущих сообщениях [28,29,30].Ранее сообщалось о присутствии 5-FQA в зеленом салате [31]. Пик 5 , который представлял исходный ион MS с m / z 473, был идентифицирован как дикаффеоилвинная кислота (DCTA) на основании его соответствия времени удерживания, УФ / видимых спектров, МС и МС 2 картины фрагментации аутентичный стандарт. Подобно предыдущим исследованиям [5, 17, 24], DCTA оказалась самой распространенной фенольной кислотой в салате-латуке. Пики 6 , 9 и 10 были идентифицированы как производные кемпферола, а именно глюкуронид кемпферола (KGR), малонилглюкозид кемпферола (KMG) и глюкуронид метилкэмпферола (MKGR), соответственно, на основе масс-спектров УФ / видимой области. данные предыдущих отчетов [5, 11, 24, 32].[M-H] — при m / z 461 и [Y o ] — при m / z 285 пика 6 были характерными свойствами KGR. Хотя лютеолин и агликон кемпферола являются изобарическими, их производные можно различить на основании данных MS n [24]. Пик 6 давал базовый пик при m / z 285 ([Yo] —) в функции высокой энергии, указывающей, что он является производным кемпферола.
Пик 5 , который представлял исходный ион MS с m / z 473, был идентифицирован как дикаффеоилвинная кислота (DCTA) на основании его соответствия времени удерживания, УФ / видимых спектров, МС и МС 2 картины фрагментации аутентичный стандарт. Подобно предыдущим исследованиям [5, 17, 24], DCTA оказалась самой распространенной фенольной кислотой в салате-латуке. Пики 6 , 9 и 10 были идентифицированы как производные кемпферола, а именно глюкуронид кемпферола (KGR), малонилглюкозид кемпферола (KMG) и глюкуронид метилкэмпферола (MKGR), соответственно, на основе масс-спектров УФ / видимой области. данные предыдущих отчетов [5, 11, 24, 32].[M-H] — при m / z 461 и [Y o ] — при m / z 285 пика 6 были характерными свойствами KGR. Хотя лютеолин и агликон кемпферола являются изобарическими, их производные можно различить на основании данных MS n [24]. Пик 6 давал базовый пик при m / z 285 ([Yo] —) в функции высокой энергии, указывающей, что он является производным кемпферола.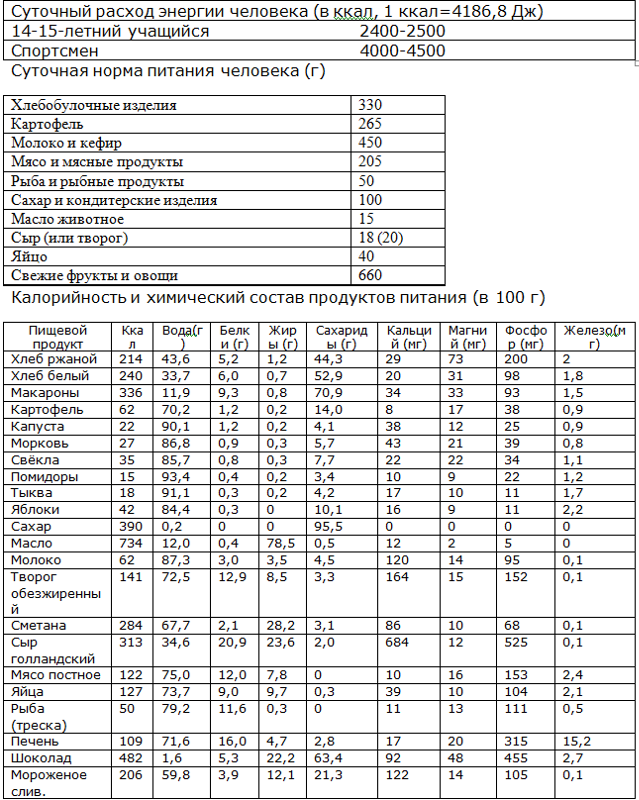 В функции низкой энергии пик 9 представлен [M-H] — при m / z 533 и базовый пик при m / z 489 из-за потери CO 2 .Функция высокой энергии дала базовый пик при m / z 285 ([Yo] —), предполагая, что это соединение является производным кемпферола. На основании данных УФ / видимого спектра и масс-спектрометрии пик 9 был идентифицирован как KMG. Глюкозиды, глюкурониды и малонилглюкозиды кемпферола уже были обнаружены в салате-латуке [5, 11, 24, 32]. Пик 10 представляет [MH] — при m / z 475 как основной пик и димерный ион при m / z 951. МС / МС дает фрагментные ионы с m / z 299, 285 и 284, которые может быть связано с потерей глюкуронидной части родительского иона, агликона кемпферола, и потерей метильной группы у иона при m / z 299, соответственно.Следовательно, этот пик был предварительно идентифицирован как глюкуронид метилкэмпферола. Метиловый конъюгат глюкуронида кемпферола может быть результатом метилирования метанолом во время процесса экстракции смесью метанол / вода / муравьиная кислота, как отмечалось в предыдущем отчете [33].
В функции низкой энергии пик 9 представлен [M-H] — при m / z 533 и базовый пик при m / z 489 из-за потери CO 2 .Функция высокой энергии дала базовый пик при m / z 285 ([Yo] —), предполагая, что это соединение является производным кемпферола. На основании данных УФ / видимого спектра и масс-спектрометрии пик 9 был идентифицирован как KMG. Глюкозиды, глюкурониды и малонилглюкозиды кемпферола уже были обнаружены в салате-латуке [5, 11, 24, 32]. Пик 10 представляет [MH] — при m / z 475 как основной пик и димерный ион при m / z 951. МС / МС дает фрагментные ионы с m / z 299, 285 и 284, которые может быть связано с потерей глюкуронидной части родительского иона, агликона кемпферола, и потерей метильной группы у иона при m / z 299, соответственно.Следовательно, этот пик был предварительно идентифицирован как глюкуронид метилкэмпферола. Метиловый конъюгат глюкуронида кемпферола может быть результатом метилирования метанолом во время процесса экстракции смесью метанол / вода / муравьиная кислота, как отмечалось в предыдущем отчете [33].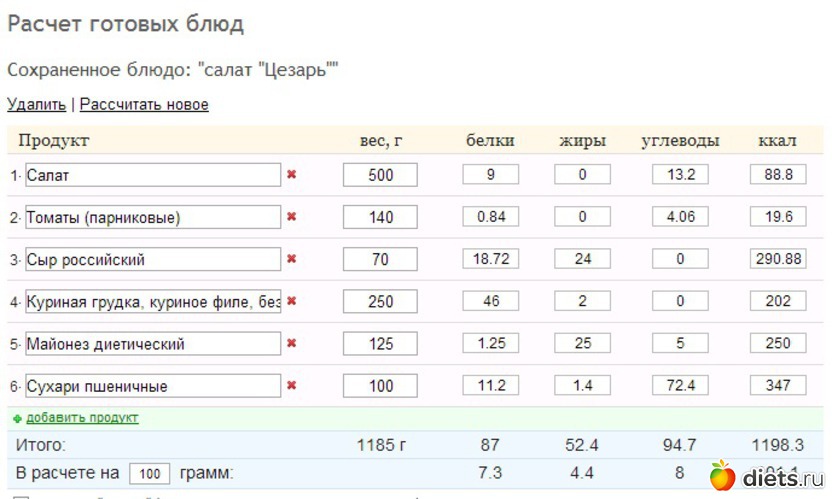 Пик 7 был однозначно идентифицирован как кверцетинмалонилглюкозид (QMG) по сравнению со временем удерживания, УФ / видимым спектром и масс-спектром аутентичного стандарта. Он представил [MH] — при m / z 549, [Y o -H] — при 300 и [MH – CO 2 ] — (базовый пик) при m / z 505, которые являются характерными свойствами QMG.Как ранее указывалось Abu-reidah et al. [34], потеря CO 2 была общей чертой соединений, представляющих малонильную группу, из-за фрагментации в источнике. Это привело к тому, что [M-H] — проявился в более низком содержании в масс-спектрах, чем ион-продукт [M-H-CO 2 ] —, как наблюдалось для пиков 7 и 9 . Идентификация QMG, которая ранее была обнаружена в салате [5, 7, 34, 35], была дополнительно подтверждена присутствием иона [2M-H] — при m / z 1099.Пик 8 , который представил [M-H] — при m / z 515, был идентифицирован как дикаффеоилхиновая кислота (DCQA).
Пик 7 был однозначно идентифицирован как кверцетинмалонилглюкозид (QMG) по сравнению со временем удерживания, УФ / видимым спектром и масс-спектром аутентичного стандарта. Он представил [MH] — при m / z 549, [Y o -H] — при 300 и [MH – CO 2 ] — (базовый пик) при m / z 505, которые являются характерными свойствами QMG.Как ранее указывалось Abu-reidah et al. [34], потеря CO 2 была общей чертой соединений, представляющих малонильную группу, из-за фрагментации в источнике. Это привело к тому, что [M-H] — проявился в более низком содержании в масс-спектрах, чем ион-продукт [M-H-CO 2 ] —, как наблюдалось для пиков 7 и 9 . Идентификация QMG, которая ранее была обнаружена в салате [5, 7, 34, 35], была дополнительно подтверждена присутствием иона [2M-H] — при m / z 1099.Пик 8 , который представил [M-H] — при m / z 515, был идентифицирован как дикаффеоилхиновая кислота (DCQA). Первый MS / MS-фрагмент DCQA произошел из-за потери одной из кофейных групп, приводящей к иону-предшественнику кофеилхиновой кислоты (при m / z 353). Последующие фрагментации представили ионы, подобные описанным выше для пиков 1 , 2 и 3 . Идентификация DCQA, о которой также сообщалось в салате [5, 7, 24, 34], дополнительно подтверждалась присутствием димерного иона при m / z 1031.
Первый MS / MS-фрагмент DCQA произошел из-за потери одной из кофейных групп, приводящей к иону-предшественнику кофеилхиновой кислоты (при m / z 353). Последующие фрагментации представили ионы, подобные описанным выше для пиков 1 , 2 и 3 . Идентификация DCQA, о которой также сообщалось в салате [5, 7, 24, 34], дополнительно подтверждалась присутствием димерного иона при m / z 1031.
Репрезентативная хроматограмма идентифицированных соединений в сортах салата. Номера пиков соответствуют идентифицированным соединениям, представленным в Таблице 1
Таблица 1 Время удерживания, максимумы поглощения в ультрафиолетовом и видимом диапазоне и данные масс-спектрометрии фенольных кислот и флавоноидов в листьях сортов салата-латукаКоличественные вариации метаболитов и антиоксидантной активности среди 22 сорта на стадии созревания и на стадии скрепления
Для исследования количественной изменчивости метаболитов в сортах салата были подготовлены репрезентативные образцы листьев для каждого сорта на стадиях созревания и скрепления. В таблице 2 показано содержание фенольной кислоты и флавоноидов в 22 сортах салата, выраженное в мг / 100 г сухого веса (DW). Калибровочные кривые были созданы с использованием аутентичных стандартов (5-CQA, DCTA, KG и QMG) для расчета результатов. Среди десяти соединений, идентифицированных с помощью UPLC-PDA-Q-TOF-HDMS, восемь из них, показывающие заметные пики, были определены количественно. Для количественного определения соединений без доступных стандартов использовались наиболее похожие соединения с учетом природы соединений и их молекулярной массы.Следовательно, 3-CQA и 5-FQA были количественно определены с использованием калибровочной кривой 5-CQA, тогда как производные кемпферола были количественно определены с использованием KG. Чтобы исследовать взаимосвязь между возрастом листьев и фенольным составом салата-латука, были количественно определены отдельные фенольные кислоты и флавоноиды в салате на стадии зрелости (товарная стадия) и стадии скрепления. Основными компонентами фенольной кислоты салата в этом исследовании были производные хинной и винной кислоты, где 5-CQA и DCTA составляли от 20 до 74% и от 15 до 56% от общего содержания суммы определенных фенольных кислот, соответственно.
В таблице 2 показано содержание фенольной кислоты и флавоноидов в 22 сортах салата, выраженное в мг / 100 г сухого веса (DW). Калибровочные кривые были созданы с использованием аутентичных стандартов (5-CQA, DCTA, KG и QMG) для расчета результатов. Среди десяти соединений, идентифицированных с помощью UPLC-PDA-Q-TOF-HDMS, восемь из них, показывающие заметные пики, были определены количественно. Для количественного определения соединений без доступных стандартов использовались наиболее похожие соединения с учетом природы соединений и их молекулярной массы.Следовательно, 3-CQA и 5-FQA были количественно определены с использованием калибровочной кривой 5-CQA, тогда как производные кемпферола были количественно определены с использованием KG. Чтобы исследовать взаимосвязь между возрастом листьев и фенольным составом салата-латука, были количественно определены отдельные фенольные кислоты и флавоноиды в салате на стадии зрелости (товарная стадия) и стадии скрепления. Основными компонентами фенольной кислоты салата в этом исследовании были производные хинной и винной кислоты, где 5-CQA и DCTA составляли от 20 до 74% и от 15 до 56% от общего содержания суммы определенных фенольных кислот, соответственно. Максимальное и минимальное содержание фенольных кислот (количественно определенная сумма отдельных фенольных кислот) в салате на стадии созревания было обнаружено у зеленых пигментированных сортов «Chunpungjeokchukmyeon» (54,6 мг / 100 г DW) и «Adam» (18,3 мг / 100 г. DW) соответственно. На стадии болтования коммерческий сорт «Томалин» с красным пигментом содержал наибольшее количество фенольной кислоты (54,6 мг / 100 г DW), в то время как сорт Pungseong с зеленым пигментом содержал наименьшее количество (15,5 мг / 100 г DW). Производные кемпферола были основными флавоноидами, обнаруженными в образцах салата.Также значительное содержание QMG измеряется в салате, особенно на стадии болтования. Сумма флавоноидов, количественно определенная в этом исследовании, колебалась от 9,2 до 25,9 мг / 100 г DW и от 14,9 до 83,0 мг / 100 г DW в образцах зрелого салата и салата на стадии болтования, соответственно. Мы заметили, что 3-CQA, KMG и MKGR были выше (до 300%) на стадии анкеровки, чем на стадии зрелости для большинства образцов.
Максимальное и минимальное содержание фенольных кислот (количественно определенная сумма отдельных фенольных кислот) в салате на стадии созревания было обнаружено у зеленых пигментированных сортов «Chunpungjeokchukmyeon» (54,6 мг / 100 г DW) и «Adam» (18,3 мг / 100 г. DW) соответственно. На стадии болтования коммерческий сорт «Томалин» с красным пигментом содержал наибольшее количество фенольной кислоты (54,6 мг / 100 г DW), в то время как сорт Pungseong с зеленым пигментом содержал наименьшее количество (15,5 мг / 100 г DW). Производные кемпферола были основными флавоноидами, обнаруженными в образцах салата.Также значительное содержание QMG измеряется в салате, особенно на стадии болтования. Сумма флавоноидов, количественно определенная в этом исследовании, колебалась от 9,2 до 25,9 мг / 100 г DW и от 14,9 до 83,0 мг / 100 г DW в образцах зрелого салата и салата на стадии болтования, соответственно. Мы заметили, что 3-CQA, KMG и MKGR были выше (до 300%) на стадии анкеровки, чем на стадии зрелости для большинства образцов.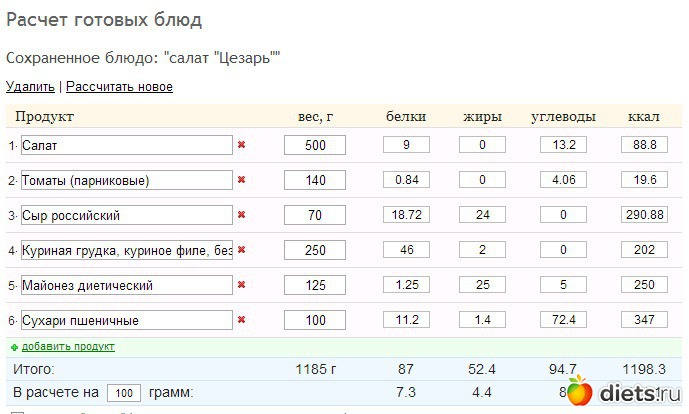 Листья салата на стадии скрепления болтами показали значительно более высокий (до 12,5 раз) QMG, чем на стадии зрелости.В отличие от других сортов, салат ромэн зеленого цвета «Чхонсан» показал в два раза меньше QMG на стадии созревания по сравнению с стадией скрепления. Однако, по крайней мере, 70% образцов показали в 4,5, 3,2 и 3,0 раза более высокое содержание 5-CQA, 5-FQA и KGR на стадии зрелости по сравнению с листьями стадии скрепления, соответственно. В нескольких образцах 1-CQA и DCQA были ниже предела количественной оценки, поэтому количественная информация не включена в этот отчет. В этом исследовании наиболее часто встречающиеся фенольные кислоты представлены хлорогеновыми кислотами и хикоровыми кислотами, в то время как флавоноиды представляют собой глюкуронид и конъюгаты глюкозида кверцетина и кемпферола, что согласуется с предыдущими сообщениями [6, 17, 24].
Листья салата на стадии скрепления болтами показали значительно более высокий (до 12,5 раз) QMG, чем на стадии зрелости.В отличие от других сортов, салат ромэн зеленого цвета «Чхонсан» показал в два раза меньше QMG на стадии созревания по сравнению с стадией скрепления. Однако, по крайней мере, 70% образцов показали в 4,5, 3,2 и 3,0 раза более высокое содержание 5-CQA, 5-FQA и KGR на стадии зрелости по сравнению с листьями стадии скрепления, соответственно. В нескольких образцах 1-CQA и DCQA были ниже предела количественной оценки, поэтому количественная информация не включена в этот отчет. В этом исследовании наиболее часто встречающиеся фенольные кислоты представлены хлорогеновыми кислотами и хикоровыми кислотами, в то время как флавоноиды представляют собой глюкуронид и конъюгаты глюкозида кверцетина и кемпферола, что согласуется с предыдущими сообщениями [6, 17, 24].
Антоцианы ответственны за красные пигменты салата-латука [1, 4]. Антоцианы демонстрируют специфические максимумы поглощения УФ / видимого света примерно при 520 нм. Идентификацию антоцианидинов (кислотно-гидролизованная форма антоцианина) проводили путем сравнения времени удерживания и УФ / видимого спектров стандартного кислотно-гидролизованного цианидина 3- O -глюкозидхлорида.Количество антоцианов определяли после кислотного гидролиза, который мог преобразовать эти пигменты в форму агликона. Цианидин был единственным основным антоцианидином, обнаруженным в этом исследовании. Содержание цианидина в сортах салата с красной и зеленой / красной пигментацией представлено на рис. 3. Однако цианидин не был обнаружен в сортах салата с зеленой пигментацией. Содержание цианидина варьировалось от 0,3 («Sunredbutter») до 9,7 мг / г DW («Tomalin») в салате зрелой стадии и от 0,5 («Superseonpung») до 10,2 мг / г DW («Jangsu») в салате стадии болтования.Статистически значимая разница ( p <0,05) в содержании цианидина наблюдалась между красными и зелеными / красными сортами салата.
Антоцианы демонстрируют специфические максимумы поглощения УФ / видимого света примерно при 520 нм. Идентификацию антоцианидинов (кислотно-гидролизованная форма антоцианина) проводили путем сравнения времени удерживания и УФ / видимого спектров стандартного кислотно-гидролизованного цианидина 3- O -глюкозидхлорида.Количество антоцианов определяли после кислотного гидролиза, который мог преобразовать эти пигменты в форму агликона. Цианидин был единственным основным антоцианидином, обнаруженным в этом исследовании. Содержание цианидина в сортах салата с красной и зеленой / красной пигментацией представлено на рис. 3. Однако цианидин не был обнаружен в сортах салата с зеленой пигментацией. Содержание цианидина варьировалось от 0,3 («Sunredbutter») до 9,7 мг / г DW («Tomalin») в салате зрелой стадии и от 0,5 («Superseonpung») до 10,2 мг / г DW («Jangsu») в салате стадии болтования.Статистически значимая разница ( p <0,05) в содержании цианидина наблюдалась между красными и зелеными / красными сортами салата. Содержание цианидина в некоторых сортах этого исследования значительно выше, чем в более раннем отчете Kim et al. [1], который зарегистрировал от 0,08 до 3,66 мг / г DW в зеленых / красных и красных листьях салата младенческого размера. Llorach et al. [5] обнаружили уровень антоцианов 0,259 и 0,456 мг / г (в пересчете на свежий вес) в образцах «красный дубовый лист» и «лолло россо» соответственно. Напротив, Перес-Лопес и др.[6], зафиксировали более высокое содержание цианидина (более 30 мг / г DW) в сорте красного пигментированного салата. Также наблюдалась значительная разница в содержании цианидина в салате-латуке на стадиях созревания и на стадии завязывания у некоторых сортов. Повышение содержания цианидина было зарегистрировано на стадии болтования для некоторых сортов салата (более 100% для «Gohong», «Sunredbutter» и «Yelpungjeokchima» и от 43 до 63% для «Jangsu», «Jeokdan» и « Jeokhagye »), тогда как снижение на 81% было продемонстрировано в Supersongpung.Для других сортов увеличение / уменьшение содержания цианидина на репродуктивной стадии составило не более 20%.
Содержание цианидина в некоторых сортах этого исследования значительно выше, чем в более раннем отчете Kim et al. [1], который зарегистрировал от 0,08 до 3,66 мг / г DW в зеленых / красных и красных листьях салата младенческого размера. Llorach et al. [5] обнаружили уровень антоцианов 0,259 и 0,456 мг / г (в пересчете на свежий вес) в образцах «красный дубовый лист» и «лолло россо» соответственно. Напротив, Перес-Лопес и др.[6], зафиксировали более высокое содержание цианидина (более 30 мг / г DW) в сорте красного пигментированного салата. Также наблюдалась значительная разница в содержании цианидина в салате-латуке на стадиях созревания и на стадии завязывания у некоторых сортов. Повышение содержания цианидина было зарегистрировано на стадии болтования для некоторых сортов салата (более 100% для «Gohong», «Sunredbutter» и «Yelpungjeokchima» и от 43 до 63% для «Jangsu», «Jeokdan» и « Jeokhagye »), тогда как снижение на 81% было продемонстрировано в Supersongpung.Для других сортов увеличение / уменьшение содержания цианидина на репродуктивной стадии составило не более 20%. Ранее сообщалось, что уровень цианидина в салате-латуке зависит от интенсивности красного цвета, генотипа, температуры и условий выращивания [4, 14, 36]. Количество накопления цианидина связано со степенью покраснения листьев, предполагая, что визуальная оценка интенсивности покраснения может помочь в качестве меры относительного количества цианидина в салате-латуке. Как сообщалось ранее [12, 37], цианидин-3-малонилглюкозид является основным антоцианом красного пигментированного салата.Помимо обеспечения сенсорных характеристик пищевых продуктов [38], цианидин-3-малонилглюкозид в красном салате, как было показано, обладает рядом полезных для здоровья свойств, таких как уменьшение фотоингибирующих и фотоокислительных повреждений [39].
Ранее сообщалось, что уровень цианидина в салате-латуке зависит от интенсивности красного цвета, генотипа, температуры и условий выращивания [4, 14, 36]. Количество накопления цианидина связано со степенью покраснения листьев, предполагая, что визуальная оценка интенсивности покраснения может помочь в качестве меры относительного количества цианидина в салате-латуке. Как сообщалось ранее [12, 37], цианидин-3-малонилглюкозид является основным антоцианом красного пигментированного салата.Помимо обеспечения сенсорных характеристик пищевых продуктов [38], цианидин-3-малонилглюкозид в красном салате, как было показано, обладает рядом полезных для здоровья свойств, таких как уменьшение фотоингибирующих и фотоокислительных повреждений [39].
Содержание цианидина в красных и зеленых / красных пигментированных сортах салата
SL — это терпеноиды C-15, которые встречаются в природе в форме углеводородов, спиртов, кетонов, альдегидов, кислот или лактонов [40]. Лактуцин и лактукопикрин, основные SL, обнаруженные в салате-латуке, вносят значительный вклад в характер горечи в сортах салата [3, 20].Два горьких сесквитерпеновых лактона (SL), лактуцин и лактукопикрин, были идентифицированы и количественно определены с помощью ВЭЖХ путем сравнения их времени удерживания и УФ / видимых спектров с соответствующими аутентичными стандартами. В таблице 3 представлены средние концентрации SLs, выраженные в микрограммах на грамм сухого веса (мкг / г DW), для 22 сортов, культивируемых на стадиях созревания и закрепления. Дисперсионный анализ (ANOVA) показал, что отдельные SL, а также общая концентрация SL показали значительные межсортовые вариации.Концентрация общих SL (сумма лактуцина и лактукопикрина) варьировалась от 11,7 («Superseonpung») до 386,7 («Sunredbutter») мкг / г DW и от 213,0 («Cheonsang») до 4101,2 («Superseonpung») мкг / г DW при стадия зрелости и стадия закрепления соответственно. Поскольку все образцы были выращены в одинаковых агрономических условиях, вариации, вероятно, будут контролироваться генетически.
Лактуцин и лактукопикрин, основные SL, обнаруженные в салате-латуке, вносят значительный вклад в характер горечи в сортах салата [3, 20].Два горьких сесквитерпеновых лактона (SL), лактуцин и лактукопикрин, были идентифицированы и количественно определены с помощью ВЭЖХ путем сравнения их времени удерживания и УФ / видимых спектров с соответствующими аутентичными стандартами. В таблице 3 представлены средние концентрации SLs, выраженные в микрограммах на грамм сухого веса (мкг / г DW), для 22 сортов, культивируемых на стадиях созревания и закрепления. Дисперсионный анализ (ANOVA) показал, что отдельные SL, а также общая концентрация SL показали значительные межсортовые вариации.Концентрация общих SL (сумма лактуцина и лактукопикрина) варьировалась от 11,7 («Superseonpung») до 386,7 («Sunredbutter») мкг / г DW и от 213,0 («Cheonsang») до 4101,2 («Superseonpung») мкг / г DW при стадия зрелости и стадия закрепления соответственно. Поскольку все образцы были выращены в одинаковых агрономических условиях, вариации, вероятно, будут контролироваться генетически. Анализ SL, включая 8-дезоксилактуцин, жаквинелин, крепидиазид B и лактузид, показал смешанные результаты среди 23 образцов Lactuca aculeata Boiss.выращивали в стандартных условиях теплицы, что позволяет предположить, что генетический фактор играет главную роль [41]. Другое исследование также показало, что концентрация лактуцина (от 2,9 до 17,2 мкг / г DW) и лактукопикрина (от 8,8 до 36,1 мкг / г DW) значительно различалась между сортами из-за цвета и морфологии листьев [3]. Было обнаружено, что содержание SL значительно варьируется в зависимости от разнообразия цикория и эндивия [42].
Анализ SL, включая 8-дезоксилактуцин, жаквинелин, крепидиазид B и лактузид, показал смешанные результаты среди 23 образцов Lactuca aculeata Boiss.выращивали в стандартных условиях теплицы, что позволяет предположить, что генетический фактор играет главную роль [41]. Другое исследование также показало, что концентрация лактуцина (от 2,9 до 17,2 мкг / г DW) и лактукопикрина (от 8,8 до 36,1 мкг / г DW) значительно различалась между сортами из-за цвета и морфологии листьев [3]. Было обнаружено, что содержание SL значительно варьируется в зависимости от разнообразия цикория и эндивия [42].
Содержание лактуцина и лактукопикрина в листьях салата на стадии скручивания выше по сравнению со зрелыми листьями салата в этом исследовании, а также в результатах предыдущих исследований [3, 15, 41].Все сорта, рассматриваемые в этом исследовании, имели более высокие концентрации лактуцина (в 1,7–118,3 раза) и лактукопикрина (в 1,7–392,7 раза) на стадии болтования по сравнению с их аналогами на зрелой стадии. Наибольшие и наименьшие изменения в содержании SL зафиксированы у сортов «Superseongpung» и «Sunredbutter» соответственно. Более высокое содержание SL на стадии скрепления по сравнению со зрелой стадией может быть связано с синтезом de novo в новых листьях, генерируемых на последней стадии, а также с измененной экспрессией SL в старых листьях.Более того, более старые ткани листа со временем могут накапливать больше SL. Литература о связи между возрастом листьев салата и SL отсутствует. Аналогичное наблюдение было получено на культурном подсолнечнике [43]. В другом исследовании общая концентрация SL прогрессивно увеличивалась по мере созревания цветков Arnica Montana от бутонов до полностью раскрытых цветков и далее увеличивалась по мере увядания лепестков [44]. Более сладкий вкус и более хрустящая текстура салата являются благоприятными сенсорными характеристиками для потребителей [45].Следовательно, сенсорные свойства, такие как горечь, частично способствуют общей приемлемости салата.
Наибольшие и наименьшие изменения в содержании SL зафиксированы у сортов «Superseongpung» и «Sunredbutter» соответственно. Более высокое содержание SL на стадии скрепления по сравнению со зрелой стадией может быть связано с синтезом de novo в новых листьях, генерируемых на последней стадии, а также с измененной экспрессией SL в старых листьях.Более того, более старые ткани листа со временем могут накапливать больше SL. Литература о связи между возрастом листьев салата и SL отсутствует. Аналогичное наблюдение было получено на культурном подсолнечнике [43]. В другом исследовании общая концентрация SL прогрессивно увеличивалась по мере созревания цветков Arnica Montana от бутонов до полностью раскрытых цветков и далее увеличивалась по мере увядания лепестков [44]. Более сладкий вкус и более хрустящая текстура салата являются благоприятными сенсорными характеристиками для потребителей [45].Следовательно, сенсорные свойства, такие как горечь, частично способствуют общей приемлемости салата. Высокое содержание SL, обнаруженное в зрелых листьях, как указано в этом исследовании, и цветущих головках растений [46] может снизить признание потребителей.
Высокое содержание SL, обнаруженное в зрелых листьях, как указано в этом исследовании, и цветущих головках растений [46] может снизить признание потребителей.
Анализ ABTS, метод, основанный на улавливании стабильного катион-радикала ABTS · + , был использован для оценки антиоксидантного потенциала экстрактов салата метанол / вода / муравьиная кислота. Анализ ABTS обеспечивает простоту работы, разнообразие и гибкость применения в различных средах для определения как гидрофильного, так и липофильного антиоксидантного потенциала пищевых экстрактов.Кроме того, поскольку ABTS имеет пики поглощения при 730 и 842 нм, он избегает помех, которые могут возникнуть из-за пигментов в экстракте салата или вторичных продуктов реакции между хромогеном и образцами [47]. Выявлены существенные различия в содержании фенольных соединений и антиоксидантной активности между зелеными, зелено-красными и красными сортами. На рис. 4 представлены возможности ABTS по улавливанию радикалов сортов салата на стадиях созревания и закрепления. Потенциал ABTS по улавливанию радикалов варьировался от 12.От 1 до 29,0 мг TE / г DW и от 15,7 до 30,3 мг TE / г DW в зрелых листьях салата на стадии созревания и стадии скручивания, соответственно. Антиоксидантная активность около 80% проанализированных сортов показала увеличение (от 2,1 до 40,9%) способности ABTS поглощать радикалы по мере созревания. В отличие от других сортов, потенциал ABTS по улавливанию радикалов красного коммерческого сорта «Superseonpung» был примерно на 50% ниже на стадии скрепления по сравнению со зрелой стадией салата. Исследования, касающиеся влияния стадии созревания на антиоксидантный потенциал салата-латука, неуловимы.Однако в случае других овощей, таких как шпинат, значения емкости поглощения радикалов кислорода (ORAC) оказались выше на стадии средней зрелости по сравнению с незрелыми и зрелыми листьями [48]. В другом исследовании пакчоя, шпината, красного салата и салата ромэн на стадии зрелой головки были выявлены более высокие значения ORAC, чем на стадии детского размера [49].
Потенциал ABTS по улавливанию радикалов варьировался от 12.От 1 до 29,0 мг TE / г DW и от 15,7 до 30,3 мг TE / г DW в зрелых листьях салата на стадии созревания и стадии скручивания, соответственно. Антиоксидантная активность около 80% проанализированных сортов показала увеличение (от 2,1 до 40,9%) способности ABTS поглощать радикалы по мере созревания. В отличие от других сортов, потенциал ABTS по улавливанию радикалов красного коммерческого сорта «Superseonpung» был примерно на 50% ниже на стадии скрепления по сравнению со зрелой стадией салата. Исследования, касающиеся влияния стадии созревания на антиоксидантный потенциал салата-латука, неуловимы.Однако в случае других овощей, таких как шпинат, значения емкости поглощения радикалов кислорода (ORAC) оказались выше на стадии средней зрелости по сравнению с незрелыми и зрелыми листьями [48]. В другом исследовании пакчоя, шпината, красного салата и салата ромэн на стадии зрелой головки были выявлены более высокие значения ORAC, чем на стадии детского размера [49]. Сорта салата с зеленой / красной и красной пигментацией превосходили по своему потенциалу поглощения радикалов ABTS по сравнению с сортами с зеленой пигментацией.Это подтверждается значительной корреляцией (коэффициент корреляции Пирсона, R = 0,811, при p <0,01) между содержанием цианидина и активностью по улавливанию радикалов ABTS, демонстрируя, что цианидин способствует антиоксидантной способности салата-латука. Более высокая антиоксидантная активность у сортов с красной и зеленой / красной пигментацией также может быть связана с их относительно большим количеством фенольных соединений. В соответствии с результатами этого исследования Pérez-López et al. [6] наблюдали, что гидрофильная антиоксидантная способность увеличивалась вместе с антоцианами для красного пигментированного салата.
Сорта салата с зеленой / красной и красной пигментацией превосходили по своему потенциалу поглощения радикалов ABTS по сравнению с сортами с зеленой пигментацией.Это подтверждается значительной корреляцией (коэффициент корреляции Пирсона, R = 0,811, при p <0,01) между содержанием цианидина и активностью по улавливанию радикалов ABTS, демонстрируя, что цианидин способствует антиоксидантной способности салата-латука. Более высокая антиоксидантная активность у сортов с красной и зеленой / красной пигментацией также может быть связана с их относительно большим количеством фенольных соединений. В соответствии с результатами этого исследования Pérez-López et al. [6] наблюдали, что гидрофильная антиоксидантная способность увеличивалась вместе с антоцианами для красного пигментированного салата.
Потенциал радикального поглощения ABTS сортов салата на стадиях созревания и насаждения
Многомерный анализ
Анализ главных компонентов (PCA) — это процедура для поиска гипотетических переменных, которые объясняют как можно большую дисперсию в многомерном наборе данных [22]. В PCA использовался многомерный анализ, чтобы различать сорта салата по цвету их листьев (зеленый, зеленый / красный и красный). Фенольные кислоты (3-CQA, 5-CQA, 5-FQA, DCTA), флавоноиды (KGR, QMG, KMG и MKGR), цианидин, SL (лактуцин и лактукопикрин) и данные об антиоксидантной активности ABTS дали три основных компонента с собственными значениями ≥ 1 корреляционной матрицы, в совокупности составляющей 84.1% и 78,0% от общей дисперсии в наборе данных по зрелым сортам салата и сортам салата в стадии болтования, соответственно (Дополнительный файл 1: Приложение S1). В зрелых сортах салата анализ Eigen нагрузок двух важных основных компонентов (PC1 и PC2) показал, что PC1 (ось X, рис. 5a) в основном вносили 5-FQA, 5-CQA и ABTS. Между тем, PC2 (ось Y, рис. 5a) в основном вносили DCTA и KGR. Первыми тремя переменными, которые вносят наибольший вклад в PC1 (ось X, рис. 5b) у зрелых листовых сортов, были 5-FQA, лактуцин и цианидин, тогда как 5-CQA, ABTS и MKGR внесли большой вклад в PC2 (ось Y, рис.
В PCA использовался многомерный анализ, чтобы различать сорта салата по цвету их листьев (зеленый, зеленый / красный и красный). Фенольные кислоты (3-CQA, 5-CQA, 5-FQA, DCTA), флавоноиды (KGR, QMG, KMG и MKGR), цианидин, SL (лактуцин и лактукопикрин) и данные об антиоксидантной активности ABTS дали три основных компонента с собственными значениями ≥ 1 корреляционной матрицы, в совокупности составляющей 84.1% и 78,0% от общей дисперсии в наборе данных по зрелым сортам салата и сортам салата в стадии болтования, соответственно (Дополнительный файл 1: Приложение S1). В зрелых сортах салата анализ Eigen нагрузок двух важных основных компонентов (PC1 и PC2) показал, что PC1 (ось X, рис. 5a) в основном вносили 5-FQA, 5-CQA и ABTS. Между тем, PC2 (ось Y, рис. 5a) в основном вносили DCTA и KGR. Первыми тремя переменными, которые вносят наибольший вклад в PC1 (ось X, рис. 5b) у зрелых листовых сортов, были 5-FQA, лактуцин и цианидин, тогда как 5-CQA, ABTS и MKGR внесли большой вклад в PC2 (ось Y, рис. .5б). График оценки и нагрузки (рис. 5) был построен на основе цвета листьев, которые были классифицированы как зеленый (7), зеленый / красный (9) и красный (6) на основе визуальной оценки в полевых условиях и в лаборатории. Как видно на рис. 5, сорта зеленого пигментированного салата были четко сгруппированы в отрицательной стороне оси X (PC1), за исключением сорта «Hacheong» зрелого салата, который находился в верхнем левом квадранте Рис. 5а. Выделение «Hacheong» из других зеленых пигментированных сортов зрелых образцов салата можно описать по значительно более высокому содержанию DCTA и KGR по сравнению с другими зелеными пигментированными сортами, которые совместно расположены в этой области загрузочного участка.Все сорта с красной пигментацией были расположены на положительной стороне PC1, за исключением листьев на стадии скручивания у сорта Superseonpung, который имел значительно более высокое содержание SL по сравнению со всеми другими изученными сортами. Зеленые / красные пигментированные сорта зрелого салата-латука были в значительной степени распределены во всех квадрантах без заметных группировок.
.5б). График оценки и нагрузки (рис. 5) был построен на основе цвета листьев, которые были классифицированы как зеленый (7), зеленый / красный (9) и красный (6) на основе визуальной оценки в полевых условиях и в лаборатории. Как видно на рис. 5, сорта зеленого пигментированного салата были четко сгруппированы в отрицательной стороне оси X (PC1), за исключением сорта «Hacheong» зрелого салата, который находился в верхнем левом квадранте Рис. 5а. Выделение «Hacheong» из других зеленых пигментированных сортов зрелых образцов салата можно описать по значительно более высокому содержанию DCTA и KGR по сравнению с другими зелеными пигментированными сортами, которые совместно расположены в этой области загрузочного участка.Все сорта с красной пигментацией были расположены на положительной стороне PC1, за исключением листьев на стадии скручивания у сорта Superseonpung, который имел значительно более высокое содержание SL по сравнению со всеми другими изученными сортами. Зеленые / красные пигментированные сорта зрелого салата-латука были в значительной степени распределены во всех квадрантах без заметных группировок.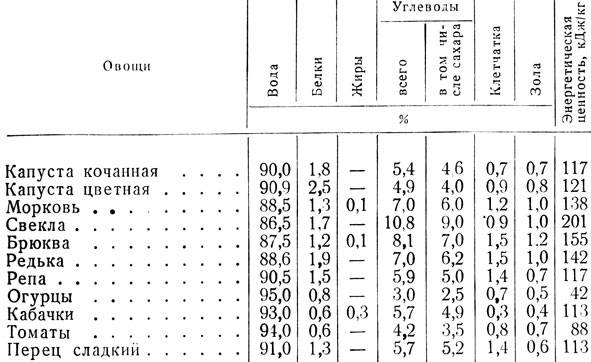 Напротив, сорта с зеленой / красной пигментацией на стадии болтования были сгруппированы в положительную сторону PC1, за исключением «Sunredbutter», который был совмещен с зелеными пигментированными сортами в нижнем левом углу квадранта.График нагрузки показал, что фенольные кислоты, флавоноиды и цианидин положительно коррелируют с ABTS, что указывает на то, что соединения вносят вклад в активность экстрактов салата по улавливанию радикалов. Образцы листьев на стадии скрепления сортов «Superseopung» и «Tomalin» отличались от других сортов, поскольку они имеют значительно более высокое содержание SL и QMG по сравнению с другими сортами, соответственно. Эти наблюдения показали, что цвет листьев может частично способствовать определенному профилю фитосоставов салата, таких как фенольные соединения и антиоксидантный потенциал.Однако корреляции между цветом листьев и содержанием SL в салате не наблюдалось.
Напротив, сорта с зеленой / красной пигментацией на стадии болтования были сгруппированы в положительную сторону PC1, за исключением «Sunredbutter», который был совмещен с зелеными пигментированными сортами в нижнем левом углу квадранта.График нагрузки показал, что фенольные кислоты, флавоноиды и цианидин положительно коррелируют с ABTS, что указывает на то, что соединения вносят вклад в активность экстрактов салата по улавливанию радикалов. Образцы листьев на стадии скрепления сортов «Superseopung» и «Tomalin» отличались от других сортов, поскольку они имеют значительно более высокое содержание SL и QMG по сравнению с другими сортами, соответственно. Эти наблюдения показали, что цвет листьев может частично способствовать определенному профилю фитосоставов салата, таких как фенольные соединения и антиоксидантный потенциал.Однако корреляции между цветом листьев и содержанием SL в салате не наблюдалось.
График загрузки и оценки 22 сортов салата на стадии зрелости ( a ) и стадии закрепления ( b ). График оценки был сгруппирован по цвету листьев (зеленый, зеленый / красный и красный). Цифры (1-22) соответствуют сортам, описанным в таблице 2
границ | Биофортификация шести сортов салата-латука (Lactuca sativa L.) йодом и селеном в сочетании с применением салициловой кислоты
Введение
Из-за широко распространенного дефицита различных минеральных компонентов в рационе человека и кормах для животных повсеместно проводятся исследования биообогащения (обогащения) культурных растений различными элементами (García-Bañuelos et al., 2014; Кшепилко и др., 2016; Przybysz et al., 2016a, b; Жук-Голашевская и др., 2016; Hegedüsová et al., 2017). Было проверено влияние йодидов и йодатов на основные физиологические и биохимические процессы в салате, выращиваемом в условиях гидропоники и беспочвенного выращивания (Blasco et al., 2010b; Voogt and Jackson, 2010; Voogt et al., 2010). Аналогичное влияние селенатов и селенитов, например, на биомассу, антиоксидантную активность, содержание вторичных метаболитов в растениях салата было также проверено (Ríos et al. , 2008, 2010).
, 2008, 2010).
Агротехнический метод биообогащения растений минеральными компонентами считается одним из самых дешевых способов сокращения их дефицита в рационе человека и кормах для животных (Zhao, McGrath, 2009; Dayod et al., 2010; Коробова, 2010). ). В случае йода (I) и селена (Se) дефицит этих элементов приходится примерно на 30 и 15% мирового населения, соответственно (White and Broadley, 2009).
За последние несколько лет был выполнен ряд исследовательских проектов, направленных на разработку агротехнических методов биофортификации (удобрения) растений отдельно с I (Cakmak, 2008; Lawson et al., 2015; Li et al., 2017) или Se (Ríos et al., 2008, 2010; Hawrylak-Nowak et al., 2015; Голубкина и др., 2017). Вопросы, касающиеся взаимодействия I и Se при одновременном удобрении этими элементами, изучены недостаточно. Оба эти элемента не являются необходимыми для правильного функционирования организмов растений. Тем не менее Se и I классифицируются в группе «полезных элементов» для растений (Kopsell and Kopsell, 2007; Medrano-Macías et al. , 2016; Gonzali et al., 2017).
, 2016; Gonzali et al., 2017).
В течение многих лет гидропонная система NFT (техника питательной пленки) использовалась в исследованиях минерального питания растений.NFT оказался совершенно правильным в исследованиях салата-латука (Rożek et al., 1989; Sady et al., 1990). Одним из преимуществ системы является простота сбора образцов листьев и корней растений для исследования без опасения их загрязнения почвой, как это часто бывает у культурных сортов, произрастающих в почве, торфе или других субстратах. Кроме того, эффективность обогащения растений минеральными компонентами (включая микроэлементы, например, I и Se) в гидропонных и / или беспочвенных культурах выше, чем в почвенных культурах (Dai et al., 2006; Риос и др., 2010; Voogt et al., 2010). Это связано с тем, что в гидропонике не происходят процессы сорбции или процессы изменения форм видообразования элементов. Эти процессы характерны для почвенной среды. Они могут ограничивать доступность минеральных компонентов для растений и, как следствие, их усвоение корнями.
В гидропонных культурах существует проблема легкого распространения грибковых и бактериальных заболеваний с питательным раствором, особенно когда культуры не оборудованы системами дезинфекции раствора.В гидропонной системе с рециркуляцией питательного раствора часто наблюдается повышенный уровень EC (электропроводность) — повышенная степень солености питательного раствора. Введение салициловой кислоты (СК) в среду в гидропонных и беспочвенных культурах может выполнять множество функций. Кислота может вызвать повышение устойчивости к абиотическим и биотическим стрессовым факторам (включая патогены) (Hayat et al., 2010). Результаты многочисленных исследований показали, что СК повышает устойчивость различных видов растений к грибковым заболеваниям (Spletzer, Enyedi, 1999; Tari et al., 2002; Mandal et al., 2009). СК, добавляемая в среду, также может увеличивать поглощение растениями минеральных компонентов, в том числе йода (Smoleń et al., 2015, 2016).
До сих пор не проводилось исследований, направленных на определение эффективности комбинированной биофортификации I и Se в различных сортах салата. Ранее такие исследования проводились, среди прочего, на ростках гороха (Jerše et al., 2018) или кольраби Osmić et al. (2017). Более того, до настоящего времени не проводилось исследований по определению различных доз экзогенной СК при поступлении и распределении I и Se (при их одновременном применении) различными сортами салата-латука.Представленное исследование восполняет информационный пробел в данной области знаний.
Ранее такие исследования проводились, среди прочего, на ростках гороха (Jerše et al., 2018) или кольраби Osmić et al. (2017). Более того, до настоящего времени не проводилось исследований по определению различных доз экзогенной СК при поступлении и распределении I и Se (при их одновременном применении) различными сортами салата-латука.Представленное исследование восполняет информационный пробел в данной области знаний.
Целью данного исследования было определение влияния экзогенной СК на процесс биообогащения растений трех ботанических разновидностей салата-латука (принадлежащих к трем типам ботанического салата) одновременно с I и Se в гидропонной системе NFT.
Материалы и методы
Растительные материалы и обработки
Двухлетние исследования были проведены в теплице с гидропонным выращиванием трех ботанических разновидностей сортов салата в системе NFT (метод питательной пленки): два сорта салата масличного (abb.BUTL) ‘Cud Voorburgu’ и ‘Zimująca’, два сорта салата айсберг (сокр. ICEL) ‘Maugli’ и ‘Królowa lata’ (все эти четыре сорта классифицируются как Lactuca sativa L. var. capitata ), как а также два сорта Lactuca sativa L. var. crispa сортов L. (сокр. REDL) «Lollo rossa» и «Redin» с небольшими красными листьями.
ICEL) ‘Maugli’ и ‘Królowa lata’ (все эти четыре сорта классифицируются как Lactuca sativa L. var. capitata ), как а также два сорта Lactuca sativa L. var. crispa сортов L. (сокр. REDL) «Lollo rossa» и «Redin» с небольшими красными листьями.
Эксперимент проводился в теплице факультета биотехнологии и садоводства Сельскохозяйственного университета в Кракове, Польша.Ежегодно семена высевали в пробки из минеральной ваты (Grodan, Rockwool B.V., Roermond, Нидерланды) в конце августа — начале сентября. Сеянцы на стадии 2 листьев помещали в отверстия (на расстоянии 25 см друг от друга) плит из пенополистирола, заполняющих грядки NFT (метод «сухой гидропоники»). Дополнительный субстрат не использовался. Теплица была оборудована пятью индивидуальными наборами NFT с емкостями для питательных растворов емкостью 1300 дм3, что облегчает выращивание салата в рециркуляционной гидропонике.
После посадки рассады растений в гидропонных системах дневная и ночная температура была установлена на 15 и 10 ° C соответственно. С начала октября до конца эксперимента естественный свет дополнялся с 5.00 до 10.00 с помощью натриевых ламп высокого давления мощностью 600 Вт.
С начала октября до конца эксперимента естественный свет дополнялся с 5.00 до 10.00 с помощью натриевых ламп высокого давления мощностью 600 Вт.
Исследование включало применение I (as KIO 3 , p.a., Avantor Performance Materials, Gliwice, Poland), Se (as Na 2 SeO 3 , puriss. P.a., Sigma-Aldrich, Сент-Луис, Миссури, США) и SA (puriss. P.a., Avantor Performance Materials) в питательный раствор для выращивания всех типов культурных сортов. Тестируемые обработки были следующими: (1) контроль (уровни следовых количеств I и Se в питательном растворе из внесенных удобрений; 30 мкг дм -3 I и 8,5 мкг дм -3 Se, соответственно), ) I + Se, (3) I + Se + 0,1 СК (0,1 мг СК дм -3 питательный раствор; т.е. 0,724 мкМ СК), (4) I + Se + 1,0 СК (1,0 мг СК ⋅ дм −3 питательный раствор; i.е., 7,24 мкМ СК) и (5) I + Se + 10,0 СК (10,0 мг СК дм -3 питательного раствора; т.е. 72,4 мкМ СК). KIO 3 использовался в дозе 5 мг I dm -3 (т. е. 39,4 мкМ I), а Na 2 SeO 3 был в дозе 0,5 мг Se dm -3 (т.е. 6,3 мкМ Se). I, Se и SA мгновенно вводились в питательный раствор, начиная с стадии 3-4 листков (формирование розетки). Эксперимент проводили по рандомизированной блочной схеме с четырьмя повторностями — по три растения в одной повторности при каждой обработке.Растения выращивали в питательном растворе с pH 5,50, EC 1,8 мСм см -1 и следующим содержанием макро- и микроэлементов (мг дм -3 ): 120 N, 40 P, 170 K, 35 Mg. , 150 Ca, 1,5 Fe, 0,55 Mn, 0,25 Zn, 0,2 B, 0,09 Cu и 0,04 Mo, что эквивалентно 8,57 мМ N, 1,29 мМ P, 4,35 мМ K, 1,44 мМ Mg, 3,74 мМ Ca, 26,9 мкМ Fe. , 10,0 мкМ Mn, 3,8 мкМ Zn, 18,5 мкМ B, 1,4 мкМ Cu и 0,4 мкМ Mo.
е. 39,4 мкМ I), а Na 2 SeO 3 был в дозе 0,5 мг Se dm -3 (т.е. 6,3 мкМ Se). I, Se и SA мгновенно вводились в питательный раствор, начиная с стадии 3-4 листков (формирование розетки). Эксперимент проводили по рандомизированной блочной схеме с четырьмя повторностями — по три растения в одной повторности при каждой обработке.Растения выращивали в питательном растворе с pH 5,50, EC 1,8 мСм см -1 и следующим содержанием макро- и микроэлементов (мг дм -3 ): 120 N, 40 P, 170 K, 35 Mg. , 150 Ca, 1,5 Fe, 0,55 Mn, 0,25 Zn, 0,2 B, 0,09 Cu и 0,04 Mo, что эквивалентно 8,57 мМ N, 1,29 мМ P, 4,35 мМ K, 1,44 мМ Mg, 3,74 мМ Ca, 26,9 мкМ Fe. , 10,0 мкМ Mn, 3,8 мкМ Zn, 18,5 мкМ B, 1,4 мкМ Cu и 0,4 мкМ Mo.
Для каждой обработки 1300 дм3 3 питательного раствора хранились в отдельных контейнерах и периодически добавлялись на плиты культивирования.Частота полива была адаптирована к стадии роста салата и погодным условиям. Растения выращивали в системе рециркуляции питательного раствора без системы дезинфекции. Растения использовали одни и те же питательные растворы на протяжении всего периода.
Растения использовали одни и те же питательные растворы на протяжении всего периода.
Сбор урожая салата с последующей оценкой веса кочана и корня и сбором образцов листьев и корней проводился в начале декабря каждого года исследования.
Анализ растений
В этой главе представлена только общая схема аналитических методов: селенометионин, селеноцистеин, пролин и SA с применением системы капиллярного электрофореза PA 800 Plus Beckman Coulter.Методы и условия, в которых проводился анализ, идентичны тем, которые использовались в нашей более ранней публикации (Smoleń et al., 2016), где представлены более подробные сведения.
Для анализов, описанных в этом подразделе, кочаны салата разрезали пополам и смешивали, чтобы получить репрезентативный образец всех листьев (старых и молодых) со всех кочанов при каждой обработке.
Образцы свежих корней и листьев салата (после промывания в дистиллированной воде) сушили при 70 ° C в лабораторной сушилке с принудительной циркуляцией воздуха и измельчали в роторной мельнице с регулируемой скоростью FRITSCH Pulverisette 14 (Идар-Оберштайн, Германия) с использованием 0. 5-мм сито. Затем образцы были проанализированы на содержание следующих элементов: I и Se в листьях и корнях с помощью метода ICP-OES.
5-мм сито. Затем образцы были проанализированы на содержание следующих элементов: I и Se в листьях и корнях с помощью метода ICP-OES.
В образцах корней и листьев также был проведен следующий анализ: селенометионин (SeMet; Acros Organics, Geel, Бельгия), селеноцистеин (SeCys; Sigma-Aldrich), пролин (Sigma-Aldrich) и SA (puriss. Pa, Avantor Performance Materials) с использованием капиллярного электрофореза через систему PA 800 Plus (Beckman Coulter).
Содержание I и Se в образцах корней и листьев после экстракции гидроксидом тетраметиламмония (TMAH) было следующим: количество 0.5 г высушенных на воздухе образцов листьев или корней, 10 см 3 бидистиллированной воды и 1 см 3 25% TMAH (Sigma-Aldrich) добавляли в 30-см пробирки 3 Falcon. После смешивания образцы инкубировали 3 ч при 90 ° C. После инкубации образцы охлаждали до температуры приблизительно 20 ° C и заполняли до 30 см 3 бидистиллированной водой. После смешивания образцы центрифугировали 15 мин при 4500 об / мин.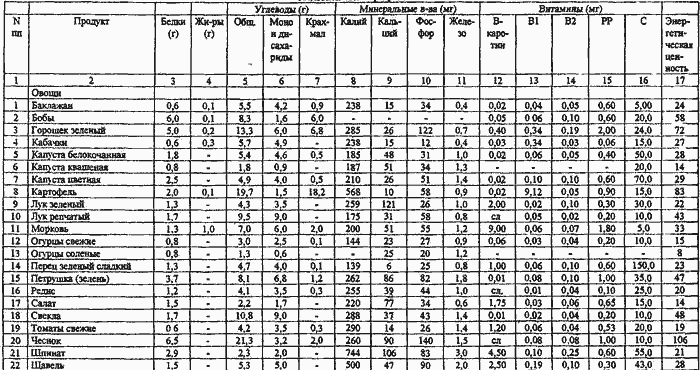 Измерения содержания I и Se с помощью спектрометра ICP-OES (Leeman Labs) проводили в супернатанте без декантации (PN-EN 15111: 2008, 2008).
Измерения содержания I и Se с помощью спектрометра ICP-OES (Leeman Labs) проводили в супернатанте без декантации (PN-EN 15111: 2008, 2008).
Анализ данных
Данные были подвергнуты дисперсионному анализу с использованием модуля двухфакторного дисперсионного анализа (обработки / это тип питательных растворов / × сорт салата) (ANOVA) программного обеспечения Statistica 10.0 PL. Было решено проверить, оказывают ли тестируемые факторы существенное влияние на биомассу салата и анализируемые параметры химического состава растений. Тест Дункана был использован для определения значимости между средними на уровне P <0.05.
Цель биофортификации
Процент рекомендуемой суточной нормы I (RDA-I) и Se (RDA-Se), поступающей из одной порции 50 г свежих листьев салата, был рассчитан с использованием результатов содержания I и Se в свежих листьях салата, а также рекомендованного суточное потребление этих двух элементов для взрослых: 150 мкг I и 55 мкг Se ежедневно (Институт медицины, 2000; Андерссон и др.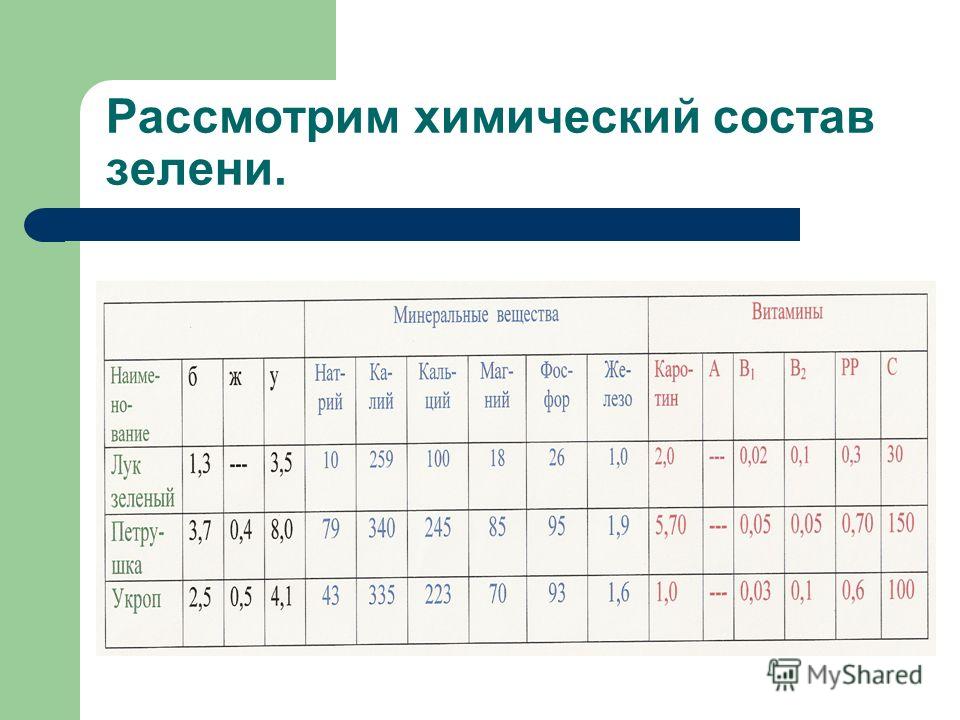 , 2007).
, 2007).
Результаты
Сорта салата показали статистически значимые и разные реакции на внесение I + Se и SA с точки зрения количества биомассы (рисунки 1A – C) и с точки зрения содержания I, Se, SeMet, SeCys. , SA и пролин в листьях и корнях (Таблицы 1–3), а также процентное содержание RDA-I и RDA-Se, а также молярное соотношение I: Se в листьях салата (Рисунки 2A – C).
Рис. 1. Влияние применения I, Se и салициловой кислоты (SA) на всю биомассу / корни + листья на растение (A) , массу одной головки салата (B) и корней на растение (С) . Средние значения, за которыми следуют разные буквы для «Обработки × Разнообразие», значительно различаются при P <0,05. Столбцы указывают на стандартную ошибку ( n = 8).
Таблица 1. Концентрации I и Se в листьях и корнях салата-латука.
Рис. 2. Процентное значение рекомендованной суточной нормы (RDA) для I (A) и Se (B) в порции свежего салата весом 50 г и соотношение молярной массы I: Se в листьях салата (С) . (A, B) Также показано увеличение столбцов на графике для контрольных растений. Средние значения, за которыми следуют разные буквы для «Обработки × Разнообразие», значительно различаются при P <0,05. Столбцы указывают на стандартную ошибку ( n = 8).
(A, B) Также показано увеличение столбцов на графике для контрольных растений. Средние значения, за которыми следуют разные буквы для «Обработки × Разнообразие», значительно различаются при P <0,05. Столбцы указывают на стандартную ошибку ( n = 8).
Биомасса салата
Среди шести сортов салата только REDL ‘Redin’ характеризовался отрицательной реакцией, выраженной в количестве биомассы, на добавление I + Se и I + Se к питательному раствору в сочетании с SA (в трех различных дозировках) (рис. 1A). –C). По сравнению с контролем внесение одного I + Se в питательный раствор и дополнительное внесение СК привело к значительному уменьшению массы кочана и корней, а также массы целых растений сорта РЕДЛ «Редин».У сорта ICEL ‘Maugli’ только совместное применение I + Se + SA в наивысшей дозе 10 мг СК дм −3 (по сравнению с остальными исследуемыми типами сред) вызывало значительное уменьшение массы кочанов. и целые растения (корни + листья / головы /).
Что касается остальных разновидностей (BUTL ‘Cud Voorburgu’, ‘Zimująca’, ‘ICEL’ Królowa lata ‘и REDL’ Lollo rossa ‘), применяемые комбинации сред с I, Se и SA по сравнению с контролем не имели влияние на количество биомассы корней, кочанов и целых растений (Рисунки 1A – C).
Содержание I, Se и полуаминокислоты в растениях салата
Добавление I и Se в питательный раствор привело к значительному увеличению содержания этих элементов в листьях и корнях всех сортов салата-латука (Таблица 1). Содержание I в листьях и корнях было примерно того же порядка, в то время как концентрация Se в корнях составляла прибл. в десять раз больше, чем в листьях. Кроме того, содержание I в листьях было от 10 до 30 раз выше, чем содержание Se.
Сравнение сортов салата с точки зрения их способности накапливать I и Se в листьях показывает, что уровни накопления йода сформированы в следующем порядке: REDL ‘Lollo rossa’> REDL ‘Redin’ = BUTL ‘Cud Voorburgu’> BUTL ‘ Zimujca ‘> ICEL’ Maugli ‘> ICEL’ Królowa lata ‘(таблица 1). В то время как содержание Se в листьях образовывалось в следующем порядке: REDL «Redin» = BUTL «Cud Voorburgu»> REDL «Lollo rossa»> ICEL «Maugli»> BUTL «Zimująca»> ICEL «Królowa lata». Таким образом, обе разновидности REDL продемонстрировали способность накапливать I и Se в наибольших количествах, в то время как ICEL «Królowa lata» характеризовалась самым низким уровнем накопления этих элементов в листьях. Важно отметить, что оба сорта REDL показали наименьшее содержание SeMet и SeCys в листьях одновременно (табл. 2).Далее, наибольшее содержание обеих этих Se-аминокислот было отмечено в сортах ICEL «Królowa lata» и BUTL «Zimująca».
В то время как содержание Se в листьях образовывалось в следующем порядке: REDL «Redin» = BUTL «Cud Voorburgu»> REDL «Lollo rossa»> ICEL «Maugli»> BUTL «Zimująca»> ICEL «Królowa lata». Таким образом, обе разновидности REDL продемонстрировали способность накапливать I и Se в наибольших количествах, в то время как ICEL «Królowa lata» характеризовалась самым низким уровнем накопления этих элементов в листьях. Важно отметить, что оба сорта REDL показали наименьшее содержание SeMet и SeCys в листьях одновременно (табл. 2).Далее, наибольшее содержание обеих этих Se-аминокислот было отмечено в сортах ICEL «Królowa lata» и BUTL «Zimująca».
Таблица 2. Концентрации селенометионина (SeMet) и селеноцистеина (SeCys) в листьях и корнях салата-латука.
Добавление СК в среду (по сравнению с выращиванием растений на питательном растворе без СК) по-разному влияло на уровень накопления I, Se, SeMet и SeCys в листьях и корнях отдельных сортов салата (таблицы 1, 2).
Влияние SA на поглощение I
Применение СК к среде (в дозе 0,1 мг mg дм -3 ) привело к значительному увеличению накопления I в листьях и корнях растений сорта ICEL ‘Królowa lata’ (таблица 1). У сортов BUTL «Zimująca» и ICEL «Maugli» только СК, внесенная в дозе 0,1 мг дм -3 , вызвала значительное увеличение степени накопления I в листьях. В листьях сорта REDL ‘Lollo rossa’ эта реакция была обнаружена после применения 10 мг SA dm -3 .Далее, у сорта BUTL ‘Cud Voorburgu’ применение СК в каждой концентрации (0,1, 1,0 и 10 мг СК дм -3 ) приводило к значительному снижению содержания I в листьях, и аналогичная реакция была обнаружена в листья растений ICEL ‘Maugli’ и REDL ‘Redin’ после внесения СК в дозах 1,0 и 10 мг СК дм −3 .
У сорта BUTL ‘Zimująca’ наблюдалось одновременное значительное увеличение степени накопления I как в листьях, так и в корнях после добавления в питательный раствор SA в 0.1 мг дм −3 доза, если сравнивать с объектом с применением только I + Se. У остальных протестированных сортов содержание I в корнях существенно не изменилось в результате внесения СК в среду.
У остальных протестированных сортов содержание I в корнях существенно не изменилось в результате внесения СК в среду.
Влияние СК на поступление селена и накопление полуаминокислоты
У всех изученных сортов салата добавление СК в питательный раствор в дозе 0,1 мг дм −3 (по сравнению с применением одного I + Se) приводило к значительному увеличению содержания Se в листьях и корнях ( Таблица 1).Однако после применения СК в двух более высоких дозах (1,0 и 10 мг СК дм -3 ) содержание Se в корнях и листьях всех сортов осталось на том же уровне или ниже, что и после внесения I + Se без SA. Единственным исключением было увеличение содержания Se в листьях сорта REDL ‘Redin’ (после применения 10 мг СК ⋅ дм -3 ) и в корнях растений сорта ICEL ‘Królowa lata’, выращиваемых на питательный раствор с добавкой 1,0 мг СК дм −3 .
Применение СК к питательному раствору в дозах 1 и 10 мг дм −3 привело к значительному увеличению содержания: (1) SeMet в листьях сортов BUTL ‘Zimująca,’ ICEL ‘Maugli’ и REDL ‘ Сорта Lollo rossa ‘, при применении дозы 10 мг SA ⋅ dm -3 , в увеличении содержания (2) SeCys в листьях обоих сортов REDL’ Redin ‘и’ Lollo rossa ‘( Таблица 2) по сравнению с выращиванием растений на питательном растворе с I + Se (без СК).
Содержание СК в растениях салата
Содержание СК в листьях и корнях отдельных сортов салата было относительно изменчивым признаком (таблица 3). В случае корней только у сорта REDL ‘Lollo rossa’ произошло значительное (по сравнению с контролем и исключительным применением I + Se) содержание СК в корнях в результате внесения СК в питательный раствор. который был в одинаковой степени во всех трех использованных дозировках SA. Что касается листьев, то только у сорта ICEL ‘Maugli’ было более чем десятикратное увеличение содержания СК в листьях растений, растущих на среде с 1.0 и 10 мг СК ⋅ дм -3 доз. Это было значительное увеличение содержания SA по сравнению с контролем и обработками I + Se и I + Se + SA 0,1 мг SA dm -3 .
Таблица 3. Влияние применения I, Se и салициловой кислоты (СК) на концентрацию СК и пролина в листьях и корнях салата-латука.
Только листья контрольных растений сортов BUTL ‘Cud Voorburgu’ и ICEL ‘Królowa lata’ показали значительно более высокое содержание СК, чем обнаруженное на средах, содержащих I + Se и добавление СК в трех разных дозах.
Листья и корни сорта BUTL ‘Zimująca’, обработанные только I + Se, содержали наибольшее количество СК, и содержание кислоты в листьях этого сорта снизилось после внесения СК в питательный раствор. Аналогичный эффект, но только в корнях, был обнаружен при удобрении I + Se в виде наибольшего накопления СК, а добавление этого соединения в питательный раствор приводило к снижению содержания СК в корнях растений. Сорта BUTL ‘Cud Voorburgu’ и ICEL ‘Królowa lata’.
Содержание пролина в растениях салата
В листьях и корнях сортов REDL «Lollo rossa» и «Redin» содержание пролина в несколько десятков раз ниже, чем в остальных сортах салата (Таблица 3). Кроме того, у этих двух сортов содержание пролина в листьях и корнях вообще не отличалось после добавления I, Se и SA в среду по сравнению с контрольными растениями.
В листьях сортов BUTL ‘Zimujca’ и ICEL ‘Królowa lata’ добавление SA в двух наивысших дозах (1.0 и 10 мг SA dm -3 ) приводили к значительному снижению содержания пролина по сравнению с применением 0,1 мг SA dm -3 .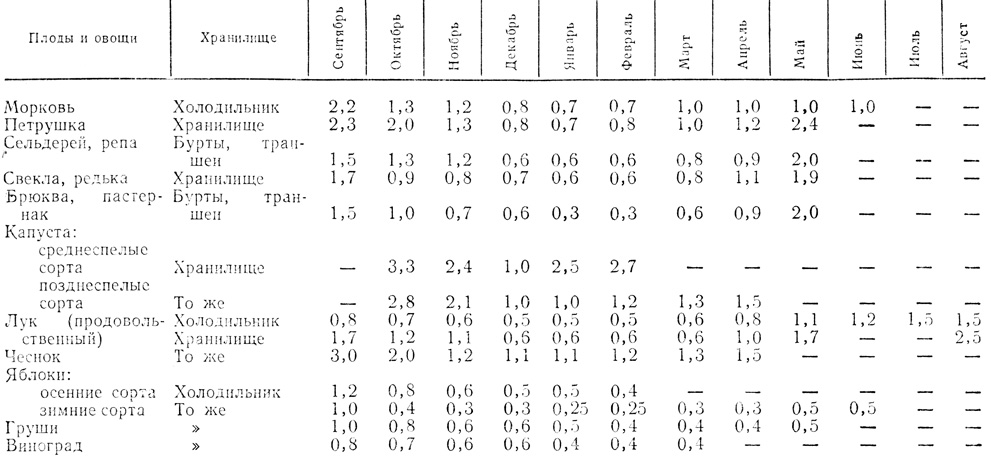
Что касается корней, то значительное увеличение содержания пролина (по сравнению с контрольными растениями и другими испытанными препаратами) было отмечено только в корнях растений сортов BUTL ‘Zimująca,’ ICEL ‘Maugli’ и ‘Królowa lata’, выращиваемых на питательный раствор с добавкой I + Se + SA 0,1 мг дм −3 .
Обсуждение
Биомасса растений и химический состав растений салата
Полученные результаты показывают, что четыре из шести сортов не показали отрицательной реакции по количеству биомассы на обогащение I и Se, а также на дополнительное внесение СК в питательный раствор.Это подтверждает, что использованные дозы I, Se и SA были безопасны для следующих сортов: BUTL ‘Cud Voorburgu’ и ‘Zimująca’, ‘ICEL’ Królowa lata ‘и REDL’ Lollo rossa ‘. Это было только в REDL’ Сорт Редин, у которого отмечена отрицательная реакция растений за счет уменьшения их биомассы на внесение в питательный раствор I + Se и I + Se + SA. Это причина того, что значительная часть этого обсуждения была посвящена этому сорту салата, а в заключительной части этой главы — снижению биомассы сорта ICEL ‘Maugli’ в ответ на I + Se + SA 10 мг. ⋅ dm −3 обработки было прокомментировано.
⋅ dm −3 обработки было прокомментировано.
Уменьшение биомассы (листья и целые растения (корни + листья / головки /) сорта REDL ‘Redin’ наблюдали в каждой из четырех обработок при выращивании на питательном растворе, обогащенном I + Se и I + Se + SA. Таким образом, вполне вероятно, что общее содержание I и Se в листьях и корнях превысило порог вредоносности для этого сорта. По нашему мнению, можно говорить только о превышении порога вредоносности, но не о пороге токсичности. обоих элементов для растений сорта РЕДЛ ‘Редин’.У растений этого сорта наблюдалось только уменьшение биомассы, но не наблюдались симптомы токсического действия, характерные для избыточного накопления I (Blasco et al., 2008, 2010a, b) и Se (Ríos et al. др., 2008, 2010; Гаврылак-Новак, 2013; Эсрингу и др., 2015). Интересно отметить, что во втором сорте группы REDL, т.е. ‘Lollo rossa’, внесение I + Se и I + Se + SA (в трех разных дозах) не привело к снижению биомассы растения.Можно предположить, что причиной различных реакций этих двух разновидностей могло быть более высокое накопление Se в разновидности «Redin», чем у разновидности «Lollo Rossa», и аналогичное меньшее накопление I.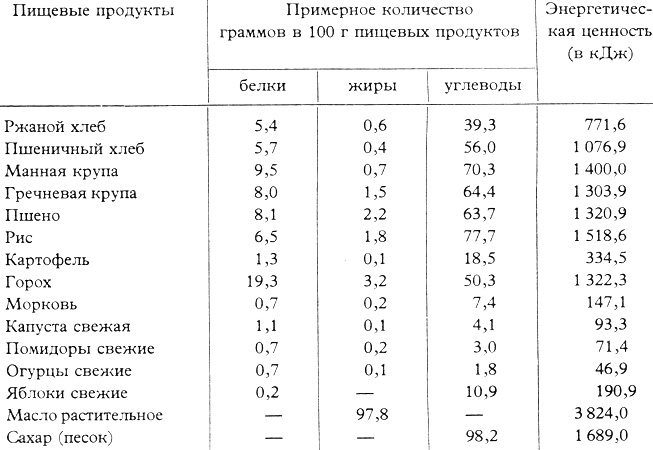 Таким образом, мы делаем вывод, что общий уровень накопления I + Se в растениях REDL «Lollo rossa» не превышал вредного уровня, как это было в случае растений REDL «Redin».
Таким образом, мы делаем вывод, что общий уровень накопления I + Se в растениях REDL «Lollo rossa» не превышал вредного уровня, как это было в случае растений REDL «Redin».
В корнях сразу после поглощения растениями иона SeO 3 2- происходит преобразование этого иона в органические соединения Se, и в основном в этой форме селен транспортируется к листьям, где он подвергается дальнейшим превращениям в другие органические соединения (Винкель и др., 2015). Усиленный синтез / накопление пролина (Hayat et al., 2012) и SA (Hayat et al., 2010) в растениях является их биохимическим ответом на возникновение абиотических или биотических стрессовых факторов. У растений сигнальная молекула, связанная со стрессовыми факторами, может быть самой SA или летучим сложным эфиром этой кислоты, то есть метилсалицилатом (MeSA). MeSA синтезируется карбоксилметилтрансферазой салициловой кислоты / SAMT / (Tieman et al., 2010). Летучий MeSA, плавающий в воздухе, служит для передачи биохимического сигнала между растениями или даже между разными листьями одного и того же растения (Taiz and Zeiger, 2010). Помимо процесса метилирования, СК у растений может претерпевать множество других преобразований, например, процесс гликозилирования, конъюгации с аминокислотами (Dempsey et al., 2011) или использоваться для образования конъюгатов сахаров с участием салициловой кислоты 3 -гидроксилаза (Zhang et al., 2013). Этот фермент приводит, например, к превращению СК в 2,3-дигидроксибензойную кислоту, то есть в дезактивированную форму СК (Hennig et al., 1993; Bartsch et al., 2010).
Помимо процесса метилирования, СК у растений может претерпевать множество других преобразований, например, процесс гликозилирования, конъюгации с аминокислотами (Dempsey et al., 2011) или использоваться для образования конъюгатов сахаров с участием салициловой кислоты 3 -гидроксилаза (Zhang et al., 2013). Этот фермент приводит, например, к превращению СК в 2,3-дигидроксибензойную кислоту, то есть в дезактивированную форму СК (Hennig et al., 1993; Bartsch et al., 2010).
В контексте изложенной выше информации интересно отметить, что у растений сорта РЕДЛ ‘Редин’ снижение продуктивности биомассы (после внесения I, Se и SA) не сопровождалось изменение содержания пролина в листьях и корнях.Однако одновременное значительное увеличение по сравнению с контролем содержания СК в листьях и корнях этого сорта наблюдалось после внесения I + Se. Кроме того, у растений REDL ‘Redin’, культивируемых на питательном растворе с наивысшей дозой экзогенной СК (10 мг СК 90 дм -3 ), самое высокое содержание СК в листьях было обнаружено внутри этого сорта, в то время как содержание СК в листьях корни были такими же, как у контрольных растений.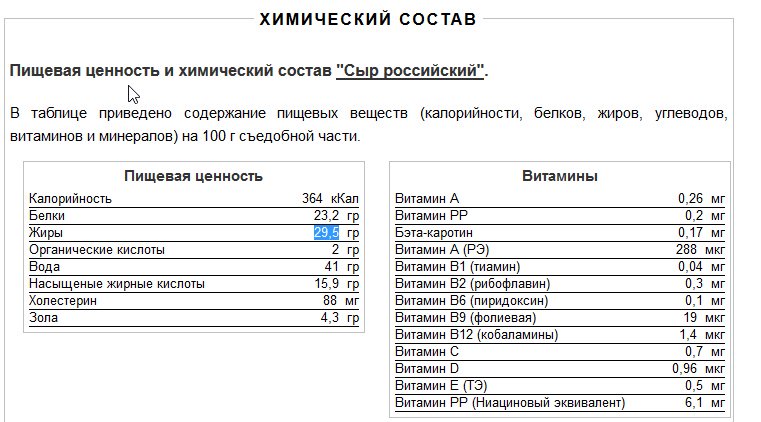
Фермент селеноцистеинметилтрансфераза (SMT) участвует в превращении SeCys в метил-SeCys.В результате потенциально токсичная Se-аминокислота (SeCys) превращается в селенометаболиты, которые менее вредны для растений (Ellis et al., 2004). Данные научных публикаций указывают на то, что у растений салата типа BUTL экзогенная СК вызывает повышение активности гена, кодирующего фермент SMT (Smoleń et al., 2016). Как следствие повышенной активности SMT, усиление процесса метилирования Se можно увидеть через синтез летучих диметилдиселенида (DMDSe) и диметилселенида (DMSe) (Winkel et al., 2015). По этой причине экзогенная СК может вызывать снижение накопления селена в листьях салата-латука BUTL-типа (Smoleń et al., 2016). В нашем исследовании взаимодействие между применением СК (дозы 1,0 и 10 против 0,1 мг СК дм -3 ) и сниженным накоплением селена в листьях двух сортов салата типа BUTL было идентично результатам исследования. Смоленя и др. (2016). Однако в случае сорта REDL «Редин» экзогенная СК вызвала усиленное накопление Se (но не I) в листьях. У этого сорта экзогенная СК, вероятно, не приводила к увеличению степени метилирования соединений Se (механизм, который мог снизить содержание Se в листьях). Фактически, мы не проводили анализа активности гена MeSA, а также уровней синтеза DMDSe и DMSe. Однако результаты определения Se-аминокислоты (низкое содержание SeMet и SeCys) показывают, что после применения экзогенной СК у обеих разновидностей типа REDL большая часть пула SeO 3 2- ионов, имеющих усваиваемые растениями, должны оставаться там в этой форме или превращаться в органические формы Se, отличные от SeMet и SeCys.Таким образом, с большой вероятностью можно предположить, что экзогенная СК может замедлять процесс метилирования DMDSe и DMSe в растениях REDL «Redin». Мы пришли к такому выводу, потому что аминокислоты SeMet и SeCys являются незаменимыми предшественниками для синтеза этих двух летучих соединений Se (Winkel et al., 2015). В связи с этим после применения SA должно происходить уменьшение количества Se, улетучивающегося в атмосферу из листьев REDL «Redin».
У этого сорта экзогенная СК, вероятно, не приводила к увеличению степени метилирования соединений Se (механизм, который мог снизить содержание Se в листьях). Фактически, мы не проводили анализа активности гена MeSA, а также уровней синтеза DMDSe и DMSe. Однако результаты определения Se-аминокислоты (низкое содержание SeMet и SeCys) показывают, что после применения экзогенной СК у обеих разновидностей типа REDL большая часть пула SeO 3 2- ионов, имеющих усваиваемые растениями, должны оставаться там в этой форме или превращаться в органические формы Se, отличные от SeMet и SeCys.Таким образом, с большой вероятностью можно предположить, что экзогенная СК может замедлять процесс метилирования DMDSe и DMSe в растениях REDL «Redin». Мы пришли к такому выводу, потому что аминокислоты SeMet и SeCys являются незаменимыми предшественниками для синтеза этих двух летучих соединений Se (Winkel et al., 2015). В связи с этим после применения SA должно происходить уменьшение количества Se, улетучивающегося в атмосферу из листьев REDL «Redin». В свете вышеизложенного, на заводах REDL ‘Redin’ должно было наблюдаться определенное падение продуктивности биомассы.Вероятно, это связано с низкой способностью этого сорта синтезировать SeMet и SeCys (и, как следствие, с более низкими уровнями синтеза DMDSe и DMSe). Это привело к повышенному уровню накопления Se после применения экзогенно более высокой SA — до уровня, превышающего порог вредоносности Se (больше Se и I) для растений сорта REDL «Редин».
В свете вышеизложенного, на заводах REDL ‘Redin’ должно было наблюдаться определенное падение продуктивности биомассы.Вероятно, это связано с низкой способностью этого сорта синтезировать SeMet и SeCys (и, как следствие, с более низкими уровнями синтеза DMDSe и DMSe). Это привело к повышенному уровню накопления Se после применения экзогенно более высокой SA — до уровня, превышающего порог вредоносности Se (больше Se и I) для растений сорта REDL «Редин».
Как уже упоминалось, СК представляет собой соединение, отвечающее за реакцию растений на абиотические и биотические стрессовые факторы (Hayat et al., 2010). Результаты нашего исследования косвенно указывают на то, что ни в одном из шести сортов салата экзогенно предоставленная СК не участвовала в защитных механизмах у растений, которые могли быть направлены на снижение степени накопления I и Se. Напротив, у изученных сортов СК (реакция, специфичная для дозы СК) приводила к повышенной степени накопления I и / или Se в листьях салата.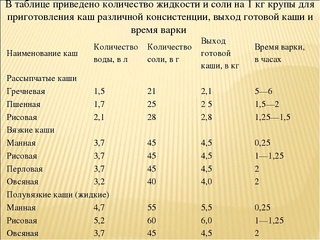 Специфическим эффектом у всех сортов был стимулирующий эффект самой низкой концентрации 0.1 мг СК ⋅ дм -3 при увеличении накопления селена в листьях и корнях. Это могло быть связано с активацией процессов, ответственных за метилирование или гидролиз СК в растениях (Hennig et al., 1993; Bartsch et al., 2010; Dempsey et al., 2011; Zhang et al., 2013), в за счет ослабления процессов, связанных с синтезом DMDSe и DMSe. Повышение эффективности или инициирование вышеупомянутых процессов, связанных с метаболизмом СК, требует больших затрат энергии.Возможно, для салата-латука процесс метилирования SA в MeSA имеет больший приоритет, чем процесс метилирования Se. На основании повышенной степени накопления селена (в результате применения СК) мы заключаем, что растения салата сорта REDL ‘Redin’, вероятно, могут вкладывать больше энергии в метилирование MeSA, чем в процессе синтеза летучих Соединения DMDSe и DMSe.
Специфическим эффектом у всех сортов был стимулирующий эффект самой низкой концентрации 0.1 мг СК ⋅ дм -3 при увеличении накопления селена в листьях и корнях. Это могло быть связано с активацией процессов, ответственных за метилирование или гидролиз СК в растениях (Hennig et al., 1993; Bartsch et al., 2010; Dempsey et al., 2011; Zhang et al., 2013), в за счет ослабления процессов, связанных с синтезом DMDSe и DMSe. Повышение эффективности или инициирование вышеупомянутых процессов, связанных с метаболизмом СК, требует больших затрат энергии.Возможно, для салата-латука процесс метилирования SA в MeSA имеет больший приоритет, чем процесс метилирования Se. На основании повышенной степени накопления селена (в результате применения СК) мы заключаем, что растения салата сорта REDL ‘Redin’, вероятно, могут вкладывать больше энергии в метилирование MeSA, чем в процессе синтеза летучих Соединения DMDSe и DMSe.
У сорта ICEL ‘Maugli’ отношения между поглощением и метаболическими путями I, Se и SA выглядят иначе, чем у разновидности REDL ‘Redin’. Обнаруженное у сорта ICEL ‘Maugli’ значительное уменьшение размеров биомасс корней и головок (только после применения I + Se + SA 10 мг ⋅ дм -3 ) должно было быть результатом совместного действия экзогенных СК на корни и одновременное накопление большого количества СК, I и Se в листьях. Наивысшее содержание СК, определенное в листьях сорта ICEL ‘Maugli’ (при обработке I + Se + SA 1.0 и I + Se + SA 10), могло быть специфической реакцией этого сорта на экзогенную SA. Эта реакция может включать усиленный синтез этого соединения в листьях и / или эффективную транспортировку экзогенной СК от корней к листьям.
Обнаруженное у сорта ICEL ‘Maugli’ значительное уменьшение размеров биомасс корней и головок (только после применения I + Se + SA 10 мг ⋅ дм -3 ) должно было быть результатом совместного действия экзогенных СК на корни и одновременное накопление большого количества СК, I и Se в листьях. Наивысшее содержание СК, определенное в листьях сорта ICEL ‘Maugli’ (при обработке I + Se + SA 1.0 и I + Se + SA 10), могло быть специфической реакцией этого сорта на экзогенную SA. Эта реакция может включать усиленный синтез этого соединения в листьях и / или эффективную транспортировку экзогенной СК от корней к листьям.
Эффект удобрения селеном может включать увеличение содержания растворимых сахаров в листьях, которые были, например, обнаружены в растениях люцерны / Medicago sativa L. / (Owusu-Sekyere et al., 2013). После применения I + Se + SA в дозе 10 мг дм -3 наибольшее содержание SeMet было обнаружено в листьях сорта ICEL ‘Maugli’. В рамках этих обработок также было самое высокое содержание сахара [сумма сахаров / глюкозы + фруктозы + сахарозы / применяемых в виде 1000 мг сахаров 100 г -1 ф. ш. of leaves], как в среднем в листьях, где app. 500–600 мг сахара ⋅ 100 г -1 сырой массы листьев по сравнению с пятью оставшимися сортами и испытанными средами (данные не представлены). По нашей оценке, большое накопление SeMet в листьях сорта ICEL ‘Maugli’ (I + Se + SA 10 мг дм -3 ) может указывать на повышенный уровень метаболизма Se-соединений, включая синтез летучих DMSe, предшественником которых является SeMet. Такая интерпретация кажется правильной, потому что растениям нужна энергия, необходимая для метаболического пути, ответственного за превращение SeMet в DMSe (Winkel et al., 2015). Опять же, в листьях салата ICEL ‘Maugli’, обработанных I + Se + SA в дозах 1,0 и 10 мг дм -3 , было отмечено заметное снижение содержания I, которое, вероятно, также было подвержено воздействию энергии потребляя процессы улетучивания (метилирования) до CH 3 I. Энергия, необходимая для метилирования соединений I и Se, вырабатывается в процессах дыхания в клетках через окисляющие сахара.
ш. of leaves], как в среднем в листьях, где app. 500–600 мг сахара ⋅ 100 г -1 сырой массы листьев по сравнению с пятью оставшимися сортами и испытанными средами (данные не представлены). По нашей оценке, большое накопление SeMet в листьях сорта ICEL ‘Maugli’ (I + Se + SA 10 мг дм -3 ) может указывать на повышенный уровень метаболизма Se-соединений, включая синтез летучих DMSe, предшественником которых является SeMet. Такая интерпретация кажется правильной, потому что растениям нужна энергия, необходимая для метаболического пути, ответственного за превращение SeMet в DMSe (Winkel et al., 2015). Опять же, в листьях салата ICEL ‘Maugli’, обработанных I + Se + SA в дозах 1,0 и 10 мг дм -3 , было отмечено заметное снижение содержания I, которое, вероятно, также было подвержено воздействию энергии потребляя процессы улетучивания (метилирования) до CH 3 I. Энергия, необходимая для метилирования соединений I и Se, вырабатывается в процессах дыхания в клетках через окисляющие сахара.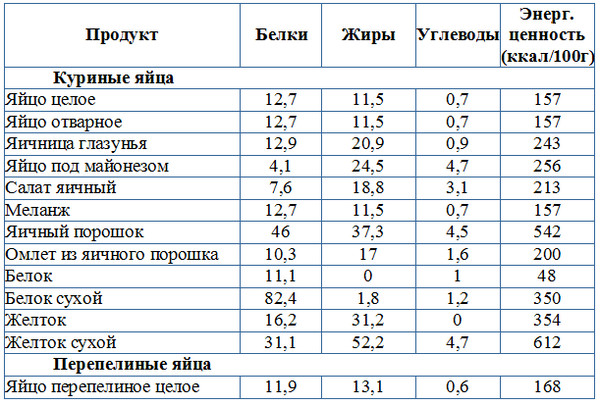 Повышенное потребление сахаров в процессах метилирования I и Se привело к недоступности этой энергии для образования биомассы растений ICEL «Маугли».Такой способ интерпретации результатов дополнительно подкрепляется тем фактом, что вместе с увеличением концентрации СК в питательном растворе было обнаружено явное падение содержания Se (а также, в определенной степени, содержания SeMet и SeCys) в корней ICEL ‘Maugli’. Это указывает на то, что экзогенная СК в дозе 10 мг дм -3 питательного раствора в растениях ICEL ‘Maugli’ оказывала максимально стимулирующее действие на процесс транспорта Se (SeO 3 −2 и / или Se-органических соединений) от корней до листьев, где они, несомненно, подверглись процессу метилирования.
Повышенное потребление сахаров в процессах метилирования I и Se привело к недоступности этой энергии для образования биомассы растений ICEL «Маугли».Такой способ интерпретации результатов дополнительно подкрепляется тем фактом, что вместе с увеличением концентрации СК в питательном растворе было обнаружено явное падение содержания Se (а также, в определенной степени, содержания SeMet и SeCys) в корней ICEL ‘Maugli’. Это указывает на то, что экзогенная СК в дозе 10 мг дм -3 питательного раствора в растениях ICEL ‘Maugli’ оказывала максимально стимулирующее действие на процесс транспорта Se (SeO 3 −2 и / или Se-органических соединений) от корней до листьев, где они, несомненно, подверглись процессу метилирования.
Уровень содержания пролина у обоих сортов в группе REDL был в несколько сотен раз ниже, чем у остальных сортов BUTL и ICEL. Таким образом, мы делаем вывод, что условия выращивания в теплице в осенне-зимний период были более оптимальными для обоих сортов типа REDL, чем для остальных четырех сортов салата (два типа BUTL и два типа ICEL).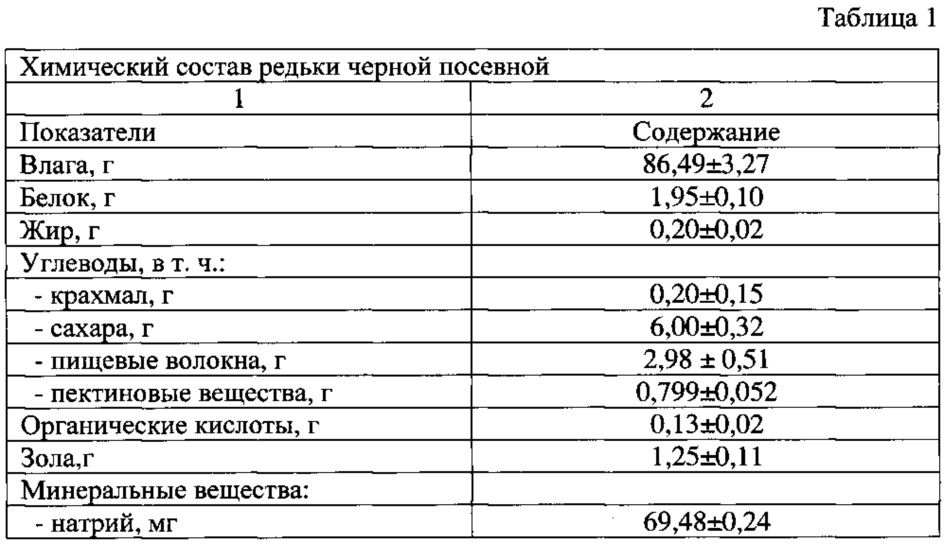 Исследования Кудела и Петржикова (2008) показали, что потенциал урожайности Lactuca sativa L.var. crispa L. салата (включая сорта «Lollo rossa» и «Redin») зависели от климатических условий и сроков выращивания (весна, лето и осень). Эти авторы обнаружили, что сорт «Редин» имеет более высокий потенциал урожайности в определенные периоды выращивания, чем «Lollo rossa». Более того, «Редин» был охарактеризован как сорт, пригодный для осеннего выращивания вне помещений. Ли и др. (2014) обнаружили, что Lactuca sativa L. var. crispa L. более устойчив к факторам абиотического стресса, чем другие ботанические сорта салата.
Исследования Кудела и Петржикова (2008) показали, что потенциал урожайности Lactuca sativa L.var. crispa L. салата (включая сорта «Lollo rossa» и «Redin») зависели от климатических условий и сроков выращивания (весна, лето и осень). Эти авторы обнаружили, что сорт «Редин» имеет более высокий потенциал урожайности в определенные периоды выращивания, чем «Lollo rossa». Более того, «Редин» был охарактеризован как сорт, пригодный для осеннего выращивания вне помещений. Ли и др. (2014) обнаружили, что Lactuca sativa L. var. crispa L. более устойчив к факторам абиотического стресса, чем другие ботанические сорта салата.
Цель биофортификации
Процент рекомендуемой суточной нормы I (RDA-I) и Se (RDA-Se), поступающей из одной порции 50 г свежих листьев салата, был рассчитан с использованием результатов содержания I и Se в свежих листьях салата, а также рекомендованного суточное потребление этих двух элементов для взрослых: 150 мкг I и 55 мкг Se ежедневно (Институт медицины, 2000; Андерссон и др. , 2007).
, 2007).
Влияние применения I, Se и SA на уровень накопления I и Se в листьях (таблица 1) было напрямую отражено в рассчитанном уровне процентного покрытия требуемых количеств RDA-I и RDA-Se. для потребителя, в результате теоретического потребления 50 г свежих листьев изученных сортов салата (рисунки 2A, B).Отношение молярного содержания I: Se в листьях определенных сортов было параметром, зависящим от добавления I, Se и SA в питательный раствор (рис. 2C).
Целью биофортификации является увеличение содержания минеральных компонентов до таких количеств, которые эффективно увеличивают возможность покрытия рациона питания потребителей определенными элементами в растениях (White and Broadley, 2009). Рекомендуемая суточная доза (RDA) для I и Se зависит от возраста и пола.Например, для взрослых RDA для этих элементов составляет 150 мкг I и 55 мкг Se. Для беременных женщин и кормящих матерей РСН увеличиваются и составляют 200–300 мкг I и 60–70 мкг Se (Институт медицины, 2000; Andersson et al. , 2007).
, 2007).
В нашем исследовании есть возможность покрыть потребности RDA-A и RDA-Se такой дозой свежих листьев салата. Результаты этих расчетов продемонстрировали, что процесс биообогащения растений I и Se эффективно увеличивает возможность удовлетворения потребностей салатом RDA-I и RDA-Se.
Среднее процентное значение покрытия требований RDA-I было самым низким у обоих сортов ICEL (121,3 и 167,7% для сортов ‘Maugli’ и ‘Królowa lata’ соответственно), а самым высоким у сорта REDL ‘Lollo rossa , то есть 348,2% RDA-I. Одновременно REDL «Lollo rossa» и BUTL «Cud Voorburgu» показали самые высокие значения индекса% RDA-Se (44,8 и 44,7% соответственно). И обе разновидности ICEL характеризовались самыми низкими значениями% RDA-Se, то есть 23,8% «Królowa lata» и 27.1% «Маугли».
Соотношение молярного содержания I: Se в пище так же важно, как и параметры RDA-I и RDA-Se. Оптимальное молярное соотношение I: Se в ежедневном потреблении пищи для людей, в зависимости от возраста и пола, рассчитанное на основе RDA-I и RDA-Se, находится в пределах 4,4–8,8: 1 (Институт медицины, 2000; Andersson et al.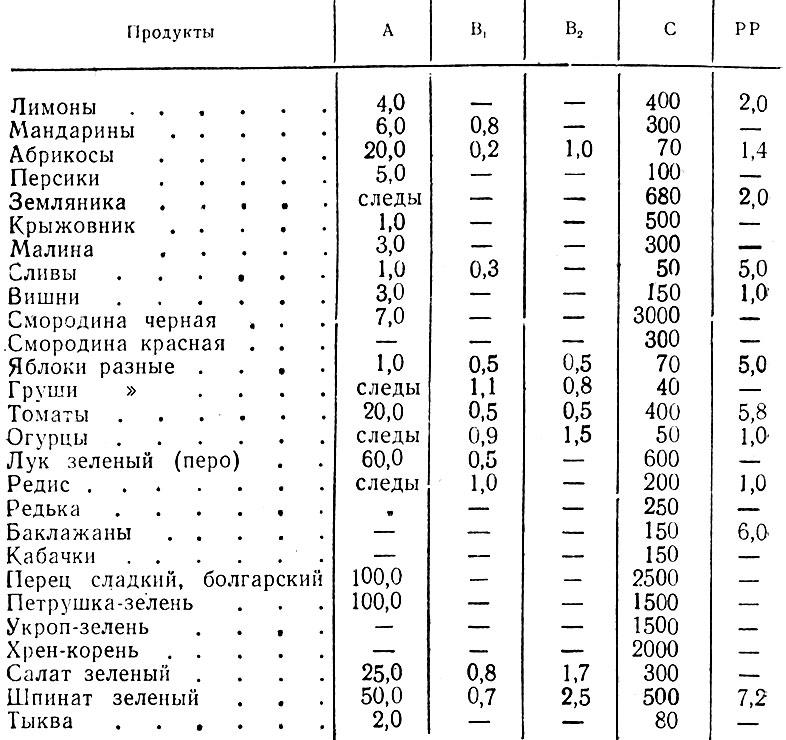 др., 2007). Необходимость поддерживать адекватное соотношение между этими элементами проистекает, например, из того факта, что I и Se выполняют очень важные роли в правильном функционировании щитовидной железы.Суть взаимосвязи I и Se в организме человека (или животного) состоит в том, что три йодтиронин дейодиназы (D1, D2 и D3) являются селензависимыми ферментами (Bianco, Kim, 2006).
др., 2007). Необходимость поддерживать адекватное соотношение между этими элементами проистекает, например, из того факта, что I и Se выполняют очень важные роли в правильном функционировании щитовидной железы.Суть взаимосвязи I и Se в организме человека (или животного) состоит в том, что три йодтиронин дейодиназы (D1, D2 и D3) являются селензависимыми ферментами (Bianco, Kim, 2006).
В нашем исследовании оба сорта ICEL, выращенные на контрольном питательном растворе (необогащенные растения), имели соотношение I: Se в листьях, т. Е. 10: 1 для сорта «Królowa lata» и 4,9: 1 для сорта «Маугли». это выгодно потребителям. Тем не менее возможности покрытия RDA-I и RDA-Se обеими разновидностями ICEL, полученными при культивировании на контрольном питательном растворе, были незначительны.
Выращивание растений на средах, обогащенных I, Se и SA, позволило получить соотношение молярного содержания I: в листьях на уровне, приближающемся к оптимальному для потребителя. Так было и с сортом ICEL ‘Maugli’, у которого соотношение I: Se в листьях в среднем составляло 7,0: 1.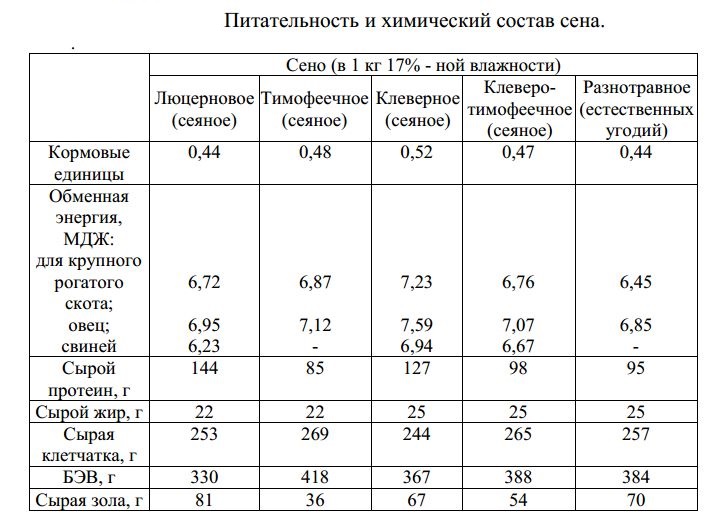 У остальных сортов рассчитанное соотношение I: Se в листьях растений, удобренных этими элементами, было несколько выше оптимума для потребителя. Средние значения для отдельных сортов были следующими: 10.9: 1 для ICEL «Królowa lata», 12,1: 1 для BUTL «Cud Voorburgu», 12,2: 1 для REDL «Redin», 14,3: 1 для BUTL «Zimująca» и 18,2: 1 для REDL «Lollo rossa»
У остальных сортов рассчитанное соотношение I: Se в листьях растений, удобренных этими элементами, было несколько выше оптимума для потребителя. Средние значения для отдельных сортов были следующими: 10.9: 1 для ICEL «Królowa lata», 12,1: 1 для BUTL «Cud Voorburgu», 12,2: 1 для REDL «Redin», 14,3: 1 для BUTL «Zimująca» и 18,2: 1 для REDL «Lollo rossa»
Заключение
Экзогенная СК, добавленная к питательному раствору, может рассматриваться как биостимулирующее соединение, улучшающее эффективность поглощения селена растениями (корнями и листьями) шести испытанных сортов салата. Этот эффект наблюдался у всех культурных сортов после применения в сочетании с самой низкой дозой СК, т.е.е., 0,1 мг дм -3 , питательный раствор.
Далее, влияние СК на накопление I в листьях было особенностью, специфичной для разновидностей и дозировки использованной кислоты. Положительное влияние всех доз экзогенной СК на увеличение степени накопления I было обнаружено у сорта ICEL ‘Królowa lata’. Самая низкая доза 0,1 мг питательного раствора SA dm -3 увеличивала эффективность I-обогащения листьев у сортов BUTL «Zimująca» и ICEL «Maugli».В случае сорта REDL «Lollo rossa» этот эффект был получен при применении СК в дозе 10 мг дм -3 . Далее, у трех сортов BUTL «Cud Voorburgu», ICEL «Maugli» и REDL «Redin» экзогенная СК способствовала уменьшению накопления I в листьях.
Самая низкая доза 0,1 мг питательного раствора SA dm -3 увеличивала эффективность I-обогащения листьев у сортов BUTL «Zimująca» и ICEL «Maugli».В случае сорта REDL «Lollo rossa» этот эффект был получен при применении СК в дозе 10 мг дм -3 . Далее, у трех сортов BUTL «Cud Voorburgu», ICEL «Maugli» и REDL «Redin» экзогенная СК способствовала уменьшению накопления I в листьях.
Мы пришли к выводу, что процессы поглощения, транспортировки и метаболизма экзогенной СК могут влиять на процесс I и Se в растениях; однако направление и интенсивность этих процессов зависят от свойств данного сорта салата.С большой долей вероятности можно предположить, что количественные изменения накопления I, Se и SA в растениях продиктованы взаимной обратной связью, основанной на ходе метаболизма I, Se и SA, характерного для каждого из испытанных сортов. салата.
В нашем исследовании аминокислота SA и пролин не были специфическими биохимическими маркерами, которые могли бы подтвердить возникновение стрессовых условий, вызванных повышенной степенью накопления I и Se в растениях салата.
Сорт REDL ‘Redin’ оказался наиболее урожайным (формируя самые большие кочаны), и одновременно он был наиболее подвержен снижению биомассы растений в результате одновременного добавления I + Se в питательный раствор, и дополнительно SA в каждой из применяемых дозировок.
Наиболее оптимальное для потребителя соотношение молярного содержания I: Se в листьях (7,0: 1) было обнаружено у сорта ICEL ‘Maugli’. У остальных сортов соотношение I: Se в листьях растений, удобренных этими элементами, было несколько выше, чем считается оптимальным для потребителя.
Авторские взносы
С.С. был руководителем проекта, соавтором метода биообогащения салата йодом и селеном с дополнительным применением салициловой кислоты, координатором эксперимента и лабораторных анализов, автором модификации методики определения йода (методом ICP- OES), а также SeMet, SeCys и пролин с помощью капиллярного электрофореза, провели анализ результатов и подготовили рукопись.И.К. являлся соавтором метода биообогащения салата йодом и селеном с дополнительным применением салициловой кислоты, проводил эксперимент с выращиванием салата, проводил химический анализ, помогал в подготовке рукописи. PK предоставил научные консультации, помощь в разработке концепции исследования и помог в подготовке рукописи. MH помог провести химический анализ образцов растений и подготовить рукопись. В.С. руководил исследованием и помогал в подготовке рукописи.
PK предоставил научные консультации, помощь в разработке концепции исследования и помог в подготовке рукописи. MH помог провести химический анализ образцов растений и подготовить рукопись. В.С. руководил исследованием и помогал в подготовке рукописи.
Финансирование
Исследование финансировалось Министерством науки и высшего образования Республики Польша.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось в отсутствие каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Благодарности
Применение салициловой кислоты с йодом в растениях было взято из нашего запатентованного решения: WS, SS и Ledwożyw-Smoleń S.Способы биообогащения овощей йодом. Патент № P.410808, Патентное ведомство Польши 07.11.2017.
Список литературы
Андерссон, М., де Бенуа, Б., Дарнтон-Хилл, И., и Деланж, Ф. (2007). Дефицит йода в Европе: постоянная проблема общественного здравоохранения.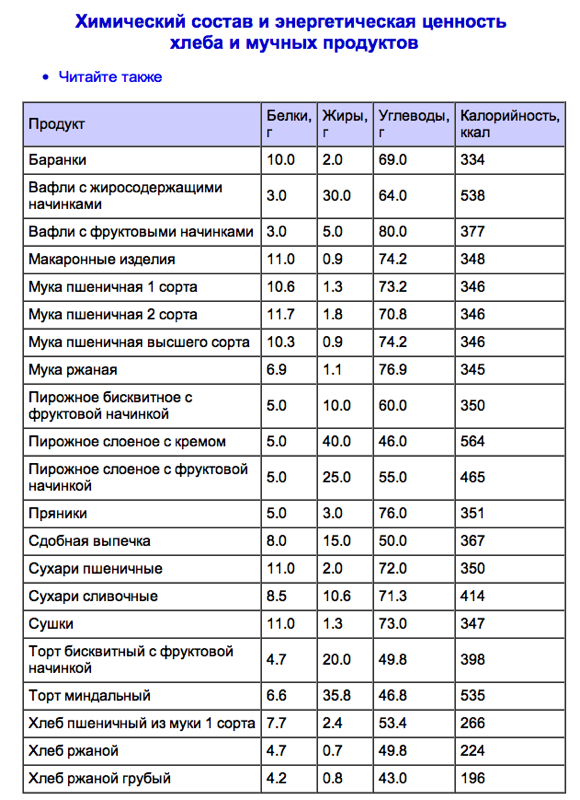 Женева: Всемирная организация здравоохранения.
Женева: Всемирная организация здравоохранения.
Google Scholar
Bartsch, M., Bednarek, P., Vivancos, P.D., Schneider, B., von Roepenack-Lahaye, E., Foyer, C.H., et al. (2010). Накопление производного изохоризматов 2, 3-дигидроксибензойного 3-O-β-D-ксилозида у Arabidopsis , устойчивости к патогенам и старения листьев. J. Biol. Chem. 285, 25654–25665. DOI: 10.1074 / jbc.M109.092569
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бласко, Б., Риос, Дж. Дж., Червилла, Л. М., Санчес-Родригес, Э., Руис, Дж. М., и Ромеро, Л. (2008). Биофортификация йодом и антиоксидантная способность салата: потенциальная польза для выращивания и здоровья человека. Ann. Appl. Биол. 152, 289–299. DOI: 10.1111 / j.1744-7348.2008.00217.x
CrossRef Полный текст | Google Scholar
Бласко, Б., Риос, Дж. Дж., Червилла, Л. М., Санчес-Родригес, Э., Рубио-Вильгельми, М. М., Розалес, М. А., и др. (2010a). Процесс фотодыхания и метаболизм азота у растений салата ( Lactuca sativa L. ) : индуцировали изменения в ответ на биофортификацию йодом. J. Регулятор роста растений. 29, 477–486. DOI: 10.1007 / s00344-010-9159-7
) : индуцировали изменения в ответ на биофортификацию йодом. J. Регулятор роста растений. 29, 477–486. DOI: 10.1007 / s00344-010-9159-7
CrossRef Полный текст | Google Scholar
Бласко, Б., Риос, Дж. Дж., Лейва, Р., Червилла, Л. М., Санчес-Родригес, Э., Rubio-Wilhelmi, M., et al. (2010b). Влияет ли биофортификация йодом на окислительный метаболизм салата-латука? Biol. Trace Elem. Res. 142, 831–842. DOI: 10.1007 / s12011-010-8816-9
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чакмак, И. (2008). Обогащение зерновых культур цинком: агрономическая или генетическая биофортификация? Почва растений 302, 1–17. DOI: 10.1007 / s11104-007-9466-3
CrossRef Полный текст | Google Scholar
Дай, Дж.Л., Чжу, Ю. Г., Хуанг, Ю. З., Чжан, М., и Сун, Дж. Л. (2006). Доступность йодида и йодата в шпинате ( Spinacia oleracea L.) по отношению к общему количеству йода в почвенном растворе. Почва растений 286, 301–308. DOI: 10.1007 / s11104-006-9139-7
DOI: 10.1007 / s11104-006-9139-7
CrossRef Полный текст | Google Scholar
Дайод, М., Тайерман, С. Д., Ли, Р. А., и Гиллихэм, М. (2010). Хранение кальция в растениях и последствия для биофортификации кальция. Protoplasma 247, 215–231.DOI: 10.1007 / s00709-010-0182-0
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Эллис, Д. Р., Сорс, Т. Г., Бранк, Д. Г., Альбрехт, К., Орсер, К., Ланер, Б. и др. (2004). Производство Se-метилселеноцистеина в трансгенных растениях, экспрессирующих селеноцистеинметилтрансферазу. BMC Plant Biol. 4: 1. DOI: 10.1186 / 1471-2229-4-1
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Эсрингу, А., Экинчи, М., Уста, С., Туран, М., Dursun, A., Ercisli, S., et al. (2015). Добавка селена влияет на рост, урожайность и накопление селена в салате-латуке ( Lactuca sativa L.). C. R. Acad. Bulg. Sci. 68, 801–810.
Google Scholar
Гарсия-Бануэлос, М. Л., Сида-Арреола, Дж. П., и Санчес, Э. (2014). Биофортификация — перспективный подход к увеличению содержания железа и цинка в основных пищевых культурах. J. Elem. 19, 865–888. DOI: 10.5601 / jelem.2014.19.3.708
Л., Сида-Арреола, Дж. П., и Санчес, Э. (2014). Биофортификация — перспективный подход к увеличению содержания железа и цинка в основных пищевых культурах. J. Elem. 19, 865–888. DOI: 10.5601 / jelem.2014.19.3.708
CrossRef Полный текст | Google Scholar
Голубкина, Н.А., Кошелева О. В., Кривенков Л. В., Добруцкая Х. Г., Надежкин С., Карузо Г. (2017). Межполые различия в росте растений, урожайности, минеральном составе и антиоксидантах шпината ( Spinacia oleracea L.) под влиянием формы селена. . Sci. Hort. 225, 350–358. DOI: 10.1016 / j.scienta.2017.07.001
CrossRef Полный текст | Google Scholar
Gonzali, S., Kiferle, C., and Perata, P. (2017). Биофортификация сельскохозяйственных культур йодом: агрономическая биофортификация, метаболическая инженерия и биодоступность йода. Curr. Мнение. Биотех. 44, 16–26. DOI: 10.1016 / j.copbio.2016.10.004
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гаврылак-Новак, Б. (2013). Сравнительное влияние селенита и селената на рост и накопление селена в растениях салата в гидропонных условиях. Регулятор роста растений 70, 149–157. DOI: 10.1007 / s10725-013-9788-5
(2013). Сравнительное влияние селенита и селената на рост и накопление селена в растениях салата в гидропонных условиях. Регулятор роста растений 70, 149–157. DOI: 10.1007 / s10725-013-9788-5
CrossRef Полный текст | Google Scholar
Гаврылак-Новак, Б., Матрашек, Р., и Погожелец, М.(2015). Двойное воздействие двух форм неорганического селена на рост, отдельные физиологические параметры и накопление макроэлементов в растениях огурца. Acta Physiol. Растение. 37:41. DOI: 10.1007 / s11738-015-1788-9
CrossRef Полный текст | Google Scholar
Хаят, К., Хакат, С., Ирфан, М., и Ахмад, А. (2010). Влияние экзогенной салициловой кислоты в изменяющейся среде: обзор. Environ. Exp. Бот. 68, 14–25. DOI: 10.1016 / j.envexpbot.2009.08.005
CrossRef Полный текст | Google Scholar
Хаят, С., Хаят, К., Алиемени, М. Н., Вани, А. С., Пичтель, Дж., И Ахмад, А. (2012). Роль пролина в меняющихся условиях Обзор. Завод Сигнал.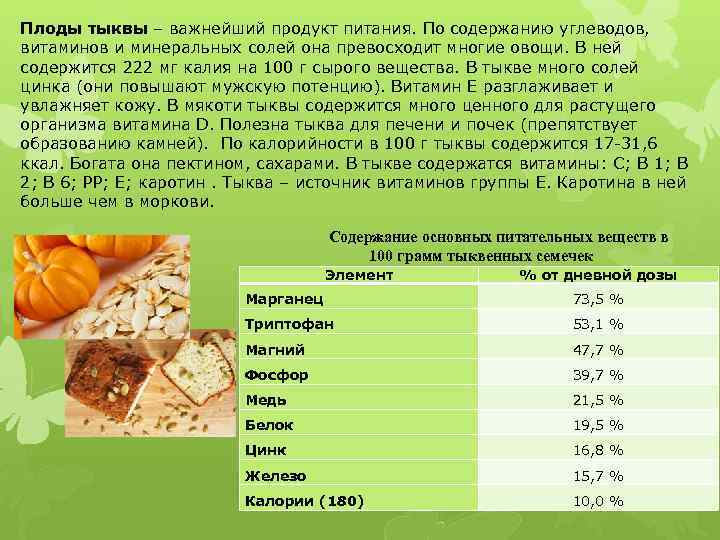 Behav. 7, 1456–1466. DOI: 10.4161 / psb.21949
Behav. 7, 1456–1466. DOI: 10.4161 / psb.21949
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Hegedüsová, A., Mezeyová, I., Hegedüs, O., Andrejiová, A., Juríková, T., Golian, M., et al. (2017). Повышение содержания и качественных показателей селена в горохе посевном ( Pisum sativum L.) после некорневой подкормки. Acta Sci. Pol. Hort. Cultus. 16, 3–17. DOI: 10.24326 / asphc.2017.6.1
CrossRef Полный текст | Google Scholar
Хенниг Дж., Малами Дж., Гринкевич Г., Индульски Дж. И Клессиг Д. Ф. (1993). Взаимное преобразование сигнала салициловой кислоты и ее глюкозида в табаке. Plant J. 4, 593–600. DOI: 10.1046 / j.1365-313X.1993.04040593.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Институт медицины (2000 г.). Нормы потребления витамина C, витамина E, селена и каротиноидов. Вашингтон, округ Колумбия: National Academies Press.
Google Scholar
Ерше, А. , Маршич, Н. К., Крофлич, А., Герм, М., Ширцель, Х., Стибиль, В. (2018). Подходит ли внекорневое обогащение растений гороха йодом и селеном для производства функционального питания? Food Chem. 267, 368–375. DOI: 10.1016 / j.foodchem.2018.02.112
, Маршич, Н. К., Крофлич, А., Герм, М., Ширцель, Х., Стибиль, В. (2018). Подходит ли внекорневое обогащение растений гороха йодом и селеном для производства функционального питания? Food Chem. 267, 368–375. DOI: 10.1016 / j.foodchem.2018.02.112
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Копселл, Д.А., Копселл Д. Э. (2007). «Селен», в справочнике по питанию растений , ред. А. В. Баркер и Д. Дж. Пилбим (Бока-Ратон, Флорида: CRC Press), 515–549.
Google Scholar
Коробова Е. (2010). Геохимические факторы почвы и ландшафта, которые способствуют пространственному распределению йода в основных компонентах окружающей среды и пищевой цепи в центральном российском плане. J. Geochem. Explor. 107, 180–192. DOI: 10.1016 / j.gexplo.2010.03.003
CrossRef Полный текст | Google Scholar
Кудела, М., и Петржикова, К. (2008). Содержание питательных веществ и урожайность отобранных сортов листового салата ( Lactuca sativa, L.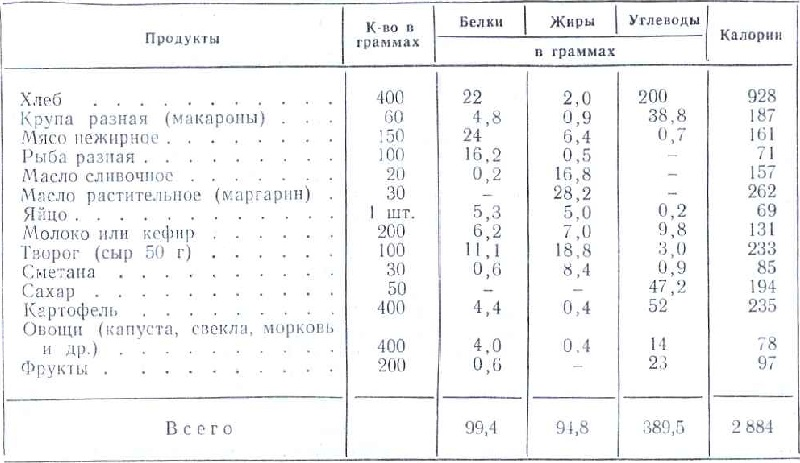 var. Crispa). Hort. Sci. 35, 99–106. DOI: 10.17221 / 3/2008-HORTSCI
var. Crispa). Hort. Sci. 35, 99–106. DOI: 10.17221 / 3/2008-HORTSCI
CrossRef Полный текст | Google Scholar
Кшепилко А., Зих-Вжик И., Молас Ю., Скварило-Беднарз Б., Свинцило А. и Сковроньска М. (2016). Влияние биофортификации йодом на отдельные биологические показатели качества проростков салата и редиса. Acta Sci.Pol. Hortorum Cultus 15, 3–16.
Google Scholar
Лоусон, П. Г., Даум, Д., Чаудерна, Р., Меузер, Х., Хертлинг, Дж. У. (2015). Сравнение почв и листовых удобрений йодом как стратегия биофортификации овощей, выращиваемых в поле. Фронт. Plant Sci. 6: 450. DOI: 10.3389 / fpls.2015.00450
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ли, К. К., Данг, Ф., Цанг, Л., Чжоу, К. Ф., и Чжоу, Д. М. (2014). Интеграция химических форм металлов и субклеточное разделение для понимания токсичности металлов в двух листьях салата ( Lactuca sativa L.) сорта. Почва растений 384, 201–212. DOI: 10.1007 / s11104-014-2194-6
DOI: 10.1007 / s11104-014-2194-6
CrossRef Полный текст | Google Scholar
Ли, Р., Ли, Д. У., Лю, Х. П., Хун, К. Л., Сонг, М. Ю., Дай, З. X. и др. (2017). Повышение содержания йода и качества плодов перца ( Capsicum annuum, L.) за счет биофортификации. Sci. Hortic. 214, 165–173. DOI: 10.1016 / j.scienta.2016.11.030
CrossRef Полный текст | Google Scholar
Мандал, С., Маллик, Н., и Митра, А. (2009). Индуцированная салициловой кислотой устойчивость к Fusarium oxysporum f. sp. lycopersici в томате. Plant Physiol. Biochem. 47, 642–649. DOI: 10.1016 / j.plaphy.2009.03.001
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Медрано-Масиас, Х., Лейя-Мартинес, П., Гонсалес-Моралес, С., Хуарес-Мальдонадо, А., и Бенавидес-Мендоса, А. (2016). Использование йода для биообогащения и повышения устойчивости сельскохозяйственных культур к стрессу и росту. Фронт.Plant Sci. 7: 1146. DOI: 10.3389 / fpls. 2016.01146
2016.01146
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Осмич, А., Голоб, А., и Герм, М. (2017). Влияние селена и йода на отдельные биохимические и морфологические характеристики проростков кольраби ( Brassica oleracea L. var. gongylodes L.). Акта Биол . Sloven. Люб. 60, 41–51.
Google Scholar
Овусу-Секьере, А., Конттури, Дж., Хаджиболанд, Р., Rahmat, S., Aliasgharzad, N., Hartikainen, H., et al. (2013). Влияние селена (Se) на углеводный обмен, клубенькообразование и рост люцерны ( Medicago sativa L.). Почва растений 373, 541–552. DOI: 10.1007 / s11104-013-1815-9
CrossRef Полный текст | Google Scholar
PN-EN 15111: 2008 (2008). Пищевые продукты — Определение микроэлементов — Определение йода с помощью ICP-MS (масс-спектрометрия с индуктивно связанной плазмой). Варшава: Польский комитет по стандартизации.
Google Scholar
Пшибыш, А., Врохна, М. , Малецка-Пшибыш, М., Гавроньска, Х. и Гавроньски, С.В. (2016a). Обогащение некоторых листовых овощей магнием. J. Elem. 21, 797–809. DOI: 10.5601 / jelem.2015.20.4.999
, Малецка-Пшибыш, М., Гавроньска, Х. и Гавроньски, С.В. (2016a). Обогащение некоторых листовых овощей магнием. J. Elem. 21, 797–809. DOI: 10.5601 / jelem.2015.20.4.999
CrossRef Полный текст | Google Scholar
Пшибыш, А., Врохна, М., Малецка-Пшибыш, М., Гавроньска, Х., Гавронски, С. В. (2016b). Влияние обогащения магнием проростков овощей на концентрацию магния, урожайность и образование АФК. J. Sci. Food Agric. 96, 3469–3476. DOI: 10.1002 / jsfa.7530
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Риос, Дж. Дж., Бласко, Б., Червилла, Л. М., Рубио-Вильгельми, М. М., Росалес, М. А., Санчес-Родригес, Э. и др. (2010). Эффективность использования азота в зависимости от различных форм и норм внесения селена в листьях салата. J. Plant Growth Reg. 29, 164–170. DOI: 10.1007 / s00344-009-9130-7
CrossRef Полный текст | Google Scholar
Риос, Дж.Дж., Росалес, М. А., Бласко, Б., Червилла, Л. М., Ромеро, Л. , и Руис, Дж. М. (2008). Биофортификация селена и индукция антиоксидантной способности растений салата. Sci. Hortic. 116, 248–255. DOI: 10.1016 / j.scienta.2008.01.008
, и Руис, Дж. М. (2008). Биофортификация селена и индукция антиоксидантной способности растений салата. Sci. Hortic. 116, 248–255. DOI: 10.1016 / j.scienta.2008.01.008
CrossRef Полный текст | Google Scholar
Рожек, С., Мычковски, Дж., Сады, В., и Войташек, Т. (1989). Влияние некоторых факторов на содержание нитратов и нитритов в листьях салата, выращенных методом питательной пленки. Я .Эффект света и регуляторы роста. Folia Hort. 1, 31–43.
Google Scholar
Сады В., Рожек С. и Грегорчик Дж. (1990). Влияние удобрений различными формами азота на урожайность и нитратный обмен в листьях тепличного салата. I. Выход и содержание отдельных компонентов в листьях салата. Folia Hort. 2, 65–76.
Google Scholar
Смолен, С., Ковальска, И., Черницка, М., Халка, М., Конска, К., и Сады, W. (2016). Биообогащение йодом и селеном с дополнительным применением салициловой кислоты влияет на урожайность, выбранные молекулярные параметры и химический состав растений салата ( Lactuca sativa L. var. capitata) . Фронт. Plant Sci. 7: 1553. DOI: 10.3389 / fpls.2016.01553
var. capitata) . Фронт. Plant Sci. 7: 1553. DOI: 10.3389 / fpls.2016.01553
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Смолен, С., Вежбиньска, Ю., Сады, В., Колтон, А., Вишневска, А., и Лиска-Скочилас, М.(2015). Биообогащение йодом с дополнительным применением салициловой кислоты влияет на урожайность и избранные параметры химического состава плодов томата ( Solanum lycopersicum L.). Sci. Hortic. 188, 89–96. DOI: 10.1016 / j.scienta.2015.03.023
CrossRef Полный текст | Google Scholar
Сплетцер, М. Э., и Эниеди, А. Дж. (1999). Салициловая кислота индуцирует устойчивость к Alternaria solani в томатах, выращенных на гидропонике. Фитопатология 89, 722–727.DOI: 10.1094 / PHYTO.1999.89.9.722
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Таиз, Л., Зейгер, Э. (2010). Физиология растений. Редвуд-Сити, Калифорния: Издательская компания Бенджамина Каммингса.
Google Scholar
Тари И., Цисар Дж., Салаи Г., Хорват Ф., Печваради А., Кисс Г. и др. (2002). Акклимация растений томатов к стрессу засоления после предварительной обработки салициловой кислотой. Acta Biol. Сегед. 46, 55–56.
Google Scholar
Тиман Д., Цейглер М., Шмельц Э., Тейлор М. Г., Рашинг С., Джонс Дж. Б. и др. (2010). Функциональный анализ метилтрансферазы салициловой кислоты томатов и ее роль в синтезе ароматизирующего летучего метилсалицилата. Plant J. 62, 113–123. DOI: 10.1111 / j.1365-313X.2010.04128.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Воогт, В., Холверда, Х. Т., и Ходабакс, Р. (2010). Биообогащение салата-латука ( Lactuca sativa L.) с йодом: влияние формы и концентрации йода в питательном растворе на рост, развитие и поглощение йода салатом, выращенным в водной культуре. J. Sci. Food Agric. 90, 906–913. DOI: 10.1002 / jsfa.3902
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Вугт, В. , и Джексон, В. А. (2010). Поглощение и распределение перхлоратов, нитратов и йода в салате-латуке ( Lactuca sativa L.) и потенциальное влияние на фоновые уровни у людей. J. Agric. Food Chem. 58, 12192–12198. DOI: 10.1021 / jf101227d
, и Джексон, В. А. (2010). Поглощение и распределение перхлоратов, нитратов и йода в салате-латуке ( Lactuca sativa L.) и потенциальное влияние на фоновые уровни у людей. J. Agric. Food Chem. 58, 12192–12198. DOI: 10.1021 / jf101227d
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Уайт, П. Дж., И Бродли, М. Р. (2009). Биообогащение сельскохозяйственных культур семью минеральными элементами, которых часто не хватает в рационе человека: железом, цинком, медью, кальцием, магнием, селеном и йодом. New Phytol. 182, 49–84. DOI: 10.1111 / j.1469-8137.2008.02738.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Винкель, Л.Х., Вринс, Б., Джонс, Г. Д., Шнайдер, Л. С., Пилон-Смитс, Э., и Бануэлос, Г. С. (2015). Цикл селена через границы раздела почва-растение-атмосфера: критический обзор. Питательные вещества 7, 4199–4239. DOI: 10.3390 / nu7064199
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чжан К., Халичке Р., Инь К., Лю К. Дж. И Ган С. С. (2013). 3-гидроксилаза салициловой кислоты регулирует продолжительность жизни листьев Arabidopsis , опосредуя катаболизм салициловой кислоты. Proc. Natl. Акад. Sci. США 110, 14807–14812. DOI: 10.1073 / pnas.1302702110
PubMed Аннотация | CrossRef Полный текст | Google Scholar
uk-Gołaszewska, K., erańska, A., Krukowska, A., and Bojarczuk, J. (2016). Биообогащение пищевой ценности пищевых продуктов из зерна Triticum durum Desf. агротехническим методом: научное обозрение. J. Elem. 21, 963–975. DOI: 10.5601 / jelem.2015.20.4.950
CrossRef Полный текст | Google Scholar
Салат Айсберг: не только вода
В течение многих лет салат айсберг не использовался в салатниках, поскольку считалось, что он не имеет питательной ценности.Айсберг обходит стороной другие, более мощные овощи, такие как салат ромэн, шпинат, красный лист салата и тому подобное. Однако многие люди предпочитают хрустящий айсберг, который до 1930-х годов назывался салат-латук с хрустящей корочкой, другим вариантам салата. Четкость айсберга уникальна. Ни один другой салат не имеет такой прохладной и приятной хрустящей корочки. Айсберг — это салат, на котором выросло мое поколение. Это не был бы салат, если бы он не начинался с айсберга.
Итак, где же «айсберг» с точки зрения питательной ценности?
Одна чашка салата айсберг содержит: 10 калорий, 0 грамм жира, 1 грамм белка, 2 грамма углеводов (1 пищевой клетчатки), 7 процентов витамина А, 3 процента витамина С, 1 процент кальция и 2 процента утюг.Он также содержит следовые количества калия, фолиевой кислоты, витамина К, магния и фосфора. Айсберг не содержит холестерина и содержит мало натрия. Конечно, это не самый питательный овощ, но если это единственный салат, который будет есть ваш ребенок, подавайте его.
Это правда, что более темная зелень содержит больше питательных веществ. Айсберг может быть приемлемым выбором в сочетании с множеством другой зелени, чтобы сделать вкусный салат. Смешивание зелени — отличный способ сохранить хрустящую корочку и питательную ценность. Листовой салат может быть немного горьким, если он слишком долго растет в земле, а иногда, когда его своевременно собирают, он все еще может сохранять некоторую горечь.Комбинируя листовой салат с айсбергом, можно уменьшить горечь, сохранив при этом другие полезные питательные вещества.
По крайней мере, салат айсберг на 96 процентов состоит из воды, так что тем из вас, кто не пьет достаточно жидкости каждый день, можно жевать кусок айсберга!
От Fine Cooking представлен рецепт салата айсберг.
Салат с рисовой лапшой по-вьетнамски
Этот салат с лапшой, наполненный ярким ароматом лайма, легким пряным жаром и нужным количеством хрустящей корочки, отлично подходит для обеда или ужина.
На 6-8 порций
½ стакана свежего сока лайма (примерно из 2-х крупных лаймов)
¼ чашки мелко нарезанной свежей кинзы
2 ст. рыбный соус *
2 ст. сахарный песок
1 ст. рубленый имбирь
3 больших измельченных зубчика чеснока
1 измельченный и очищенный от семян тайский чили
8 унций. сушеная рисовая лапша (толщиной около дюйма)
8 чашек тонко нарезанного салата айсберг (с 1 большой головки)
1 большая морковь, нарезанная лентами (используйте овощечистку)
1 большой огурец, очищенный, если хотите, нарезанный кубиками ½ дюйма
5 средних редис, тонко нарезанных
1 стакан свежих листьев мяты
½ стакана соленого арахиса, крупно нарезанного
Направление:
Заправка: В небольшой миске смешайте сок лайма, кинзу, рыбный соус, сахар, имбирь, чеснок и чили и оставьте на 10 минут.
Тем временем доведите до кипения большую кастрюлю с хорошо подсоленной водой. Добавьте рисовую лапшу и сразу же перемешайте. Готовьте лапшу, часто помешивая, примерно 4 минуты, пока она не станет мягкой. Слейте воду и промойте лапшу холодной водой, пока она не остынет на ощупь.
В большой салатнице смешайте лапшу с салатом, морковью, огурцом, редисом и листьями мяты. Перемешайте салат с заправкой и украсьте арахисом.
Информация о пищевой ценности:
На порцию: 200 калорий, 5 граммов жиров, 5 граммов белка, 36 граммов углеводов, 3 грамма пищевых волокон.
Примечание: Рыбный соус — это очень мощная азиатская приправа, сделанная из жидкости из соленой, ферментированной рыбы — отсюда и запах. Рыбный соус имеет очень концентрированный вкус и, как и паста из анчоусов, при использовании в небольших количествах придает продуктам соленый вкус, который трудно скопировать.
Джорджия Кларк-Альберт — зарегистрированный диетолог и дополнительный инструктор по питанию в Общественном колледже Восточного Мэна, проживающая в Афинах. Прочтите ее колонки и задайте вопросы по телефону bangordailynews.com или напишите ей по телефону [email protected] .
компонентов эфирного масла семян и свежих листьев салата садового (Lactuca sativa L.), выращенного в Исфахане, Иран
[1] Мозаффарян В. Идентификация лекарственных и ароматических растений Ирана. Тегеран: Фарханг-э-Моазер, 2015.
[2] Rhazes M . Аль-Хави. Vol. 20. Afsharypuor, S. Trans. Тегеран: Академия медицинских наук Ирана, 2005.
[3] Bin Sina H . АЛЬ-канун фи-аль-тиб. Нью-Дели: Хамдард Нагар, 1987.
[4] Агили Алави Хорасани Ширази MH.Махзан аль ‘Адвиех. Тегеран: Рахе Камаль, 2008.
[5] Тутин Т.Г., Хейвуд В.Х., Берджес Н.А., Мур Д.М., Валентайн Д.Х., Уолтерс С.М., Уэбб Д.А. Flora Europaea. Лондон: Издательство Кембриджского университета, 1984.
[6] Адамс Р.П.Идентификация компонентов эфирных масел методом газовой хроматографии / квадрапольной масс-спектроскопии . Иллинойс: издательство Allured, 2004.
[7] Любек А.Дж., Саттон Д.Л. Индексы удерживания Коваца отобранных углеводородов через C 10 на капиллярах из плавленого кварца со связанной фазой. J High Resolut Chromatogr Chromatogr Commun. 1983; 6 (6): 328-332.
[8] Flavornet: индексы удержания Kovats. [Проверено в 2017 г.]. Доступно по адресу: http://www.flavornet.org/ f_kovats.html.
[9] Национальный институт стандартов и технологий.NIST . [дата обращения: 2107]. Доступно по адресу: https://www.nist.gov/programs-projects/nist-chemistry-webbook.
[10] Кениг Ф., Саймонс Дж. Х., Крич Д., Коуэн Дж. П., Вентура Г. Т., Ребейн-Халили Т. Структура и распределение разветвленных алифатических алканов с четвертичными атомами углерода в сеноманских и туронских черных сланцах холмов Паскиа (Саскачеван, Канада). Org Geochem. 2005; 36 (1): 117-138.
[11] Miyazawa M, Nagai S, Oshima T. Летучие компоненты соломы Oryza sativa L. J Oleo Sci. 2008; 57 (3): 139-143.
[12] Яйли Н., Яс А., Гулец С., Уста А., Колайли С., Чоскунчелеби К., Караоглу С.Состав и антимикробная активность эфирных масел Centaurea sessilis и Centaurea armena . Фитохимия. 2005; 66 (14): 1741-1745.
[13] Кан WY, Ji ZQ, Wang JM. Состав эфирного масла Adiantum flabellulatum . Chem Nat Compd. 2009; 45 (4): 575-577.
[14] Афшарипуор С., Джамали М. Летучие компоненты надземных частей цветения, плодов и корней Cardaria draba L. J Essent Oil Res. 2006; 18 (6): 674-675.
[15] Харборн Дж.Б.Фитохимические методы. 2 -й изд. Лондон: Чепмен и Холл, 1984.
[16] Сюй Ф., Ван Ц., Айса Х.А. Анализ эфирного масла, экстрагированного из семян Lactuca sativa , произрастающих в Синьцзяне, методом ГХ-МС. Чжун Яо Кай. 2011; 34 (12): 1887–1891.
[17] Кришнамурти Р.Г., Смаус Т.Х., Мукхерджи Б.Д., Редди Б.Р., Чанг СС.Идентификация 2-пентилфурана в жирах и маслах и его связь с ароматом реверсии соевого масла. J Food Sci. 1967; 32 (4): 372-374.
[18] Аль-Номаани RSS, Хоссейн М.А., Вели А.М., Аль-Риями К., Аль-Сабахи Дж. Н.. Химический состав эфирных масел in vitro антиоксидантная активность сырых экстрактов свежих и сухих листьев лекарственного растения Lactuca sativa L.уроженец султаната Оман. Азиатский Пак Дж. Троп Биомед. 2013; 3 (5): 353-357.
[19] Эльшаркави Э., Альшатли М. Противораковая активность Lactuca serriolla , растущего в условиях сухой пустыни в северном регионе Саудовской Аравии.
